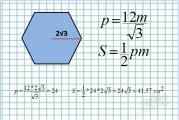
Classement des algues. Structure, reproduction des algues vertes et brunes. La valeur des algues dans l'économie nationale et la médecine. La signification des algues : classification des algues dans le dictionnaire Collier Une classification systématique des algues et des mousses
Les plantes aquatiques sont divisées en supérieur (Cormobionta) et inférieur (Thallobionta). Ces derniers comprennent tous les types d'algues. Ils sont l'un des plus anciens représentants de la flore. Leur principale caractéristique est la reproduction des spores, et la particularité réside dans la capacité de s'adapter à diverses conditions. Il existe des types d'algues qui peuvent vivre dans n'importe quelle eau : salée, fraîche, sale, propre. Mais pour les aquariophiles, ils deviennent un gros problème, surtout dans le cas de leur croissance violente.
Il existe des types d'algues qui peuvent vivre dans n'importe quelle eau : salée, fraîche, sale, propre.
Caractéristique principale
Selon les espèces d'algues, certaines sont attachées aux surfaces sous-marines, tandis que d'autres vivent librement dans l'eau. Les cultures peuvent ne contenir que du pigment vert, mais il existe des espèces avec des pigments différents. Ils colorent les algues en rose, bleu, violet, rouge et presque noir.
Les processus biologiques se produisant dans l'aquarium sont à la base de l'apparition indépendante des algues. Ils sont introduits lorsque les poissons sont nourris avec des aliments vivants ou des plantes aquatiques nouvellement acquises.
Certaines algues ressemblent à un bouquet pelucheux, d'autres ressemblent à un tapis étalé et d'autres ressemblent à une membrane muqueuse. Il existe des cultures plates, thalles, ramifiées et filamenteuses. Contrairement aux plantes supérieures, elles n'ont pas de racines, de tiges ou de feuilles. Leur forme, leur structure et leur taille sont variées. Certaines espèces ne peuvent être vues qu'au microscope. Dans le milieu naturel, les plantes atteignent plusieurs mètres de longueur.
Classement des algues
Chaque espèce a ses propres exigences pour l'environnement dans lequel elle pousse - à la température du liquide, à l'intensité et à la durée de l'éclairage. Un facteur important est la composition chimique de l'eau.
Le déséquilibre des algues dans l'aquarium indique l'apparition de conditions défavorables dans celui-ci. Une augmentation excessive de ceux-ci dans le réservoir aggrave la qualité de l'eau, ce qui nuit à la santé des habitants de l'aquarium. Une infestation d'algues peut être causée par :
- Éclairage d'aquarium non réglementé. C'est un manque d'heures de clarté ou son excès.
- Excès de matières organiques dans le contenant. Ils peuvent être sous forme de restes de nourriture, morts plantes d'aquarium, eaux usées de poisson.
- décomposition de la matière organique. L'apparition de nitrites et d'ammoniac dans l'aquarium.
Après avoir identifié quel facteur est à l'origine de l'apparition des cultures, il est nécessaire de l'éliminer ou de le minimiser autant que possible.
Le déséquilibre des algues dans l'aquarium indique l'apparition de conditions défavorables dans celui-ci.
Les algues sont divisées en 12 types. L'aquarium se caractérise le plus souvent par la présence de trois principaux types de cultures.
Leur présence est prévisible là où il y a de l'eau, de la lumière et des nutriments.
Groupe vert
C'est le groupe de plantes le plus courant et le plus diversifié en termes de structure et de forme, qui compte environ 7 000 espèces. Ils se présentent sous des formes non cellulaires, unicellulaires et multicellulaires. Les algues forment des colonies sur le verre ou le sol.
Leur particularité est que presque toutes les cultures apparaissent à la suite d'un éclairage excessif. Ils ont une couleur verte, malgré la teneur en eux, en plus de la chlorophylle verte, d'un pigment jaune. Les algues colorent le liquide en vert ou en vert brique.
Il existe des espèces marines et d'eau douce. Noms des algues présentes dans l'aquarium :
La principale raison de l'apparition de la plupart des espèces d'algues vertes est un éclairage excessif. Ainsi, lorsque l'équilibre biologique est rétabli, ce problème peut rapidement disparaître.
Plantes diatomées (brunes)
Si le liquide contenu dans le récipient doit être changé fréquemment, car il devient rapidement trouble, - algues brunes dedans. Cela gâche non seulement l'intérieur de l'aquarium, mais cause également des inconvénients à ses habitants. Ce sont des organismes microscopiques unicellulaires qui se multiplient rapidement et créent un revêtement visqueux sur les feuilles des plantes d'aquarium et des verres de réservoir. Ils vivent seuls ou en colonies sous la forme d'un ruban, d'un fil, d'une chaîne, d'un film, d'un buisson.
Sur le stade initial l'apparition de la plaque dans le récipient, elle s'enlève facilement et, dans les cas avancés, elle devient multicouche et il peut être difficile de s'en débarrasser. Les plantes brunes ne nuisent pas aux animaux d'aquarium, mais elles sont dangereuses pour les plantes d'aquarium. La plaque sur les cultures interfère avec la photosynthèse, ce qui entraîne leur mort.
La reproduction des diatomées est réalisée par division. Les cellules végétales ont une coque dure avec une composition de silice. Leurs dimensions sont au moins 0,75 µm, au maximum 1500 µm. Cette culture est facile à distinguer par la coquille sous forme de points, de chambres, de traits, de bords disposés avec une régularité géométrique.
Navicula vit presque partout, démarre au printemps et en automne.
Dans la nature, environ 25 000 variétés de cultures brunes. On trouve le plus souvent dans des conteneurs :
- Navicule. Ce genre compte environ 1 000 espèces d'algues. Planté en pots au printemps et en automne. La méthode de reproduction est la division cellulaire. Les cellules ont une forme, une structure de coque et une structure différentes. Ils servent de nourriture aux habitants de l'aquarium et eux-mêmes mangent phototrophiquement.
- Pinnulaire. Le début de l'automne et l'été sont les périodes d'apparition de ce genre. À la suite de la division cellulaire, chacun reçoit un feuillet de la cellule mère. Les cellules individuelles sont rarement connectées en rubans. Environ 80 espèces de ces algues sont connues.
- Cymbelle. Le genre est une seule cellule libre, qui est parfois attachée au substrat par une tige muqueuse. De plus, ils peuvent être enfermés dans des tubes gélatineux.
Les algues brunes se développent dans les réservoirs où l'eau ne change pas dans le temps ou l'éclairage est faible. Leur distribution est affectée par la population dense de l'aquarium, un grand nombre de matières organiques, filtre obstrué.
Rouge ou "cramoisi"
Les algues rouges, ou algues violettes, sont une petite espèce de cultures, la grande majorité sont multicellulaires, comptant jusqu'à 200 variétés. Tous les violets sont divisés en 2 classes, chacune contenant 6 commandes. Ils s'installent sur les tiges et les extrémités des feuilles des plantes d'aquarium, des pierres, poussent rapidement et se multiplient intensément.
La raison de l'apparition de ce type de plante est un excès de matière organique dans l'eau, un éclairage mal installé ou une surpopulation dans le réservoir. Ces cultures représentent un danger pour ses habitants, elles doivent donc être détruites en temps opportun.
Le cramoisi, selon la combinaison de pigments, change de couleur du rouge vif au vert bleuâtre et au jaune, et ceux d'eau douce sont généralement verts, bleus ou noir brunâtre. Une caractéristique des plantes est leur cycle de développement complexe. En règle générale, ces cultures poussent attachées à d'autres plantes, pierres, réservoirs. Vous pouvez trouver des colonies de cultures sous forme de dépôts muqueux.
Les algues rouges, ou algues violettes, sont une petite espèce de cultures, la grande majorité sont multicellulaires, comptant jusqu'à 200 variétés.
Pour les aquariophiles, il existe deux types de catastrophe :
- Barbe noire. Au stade initial, il s'agit d'un seul buisson noir concentré en un seul endroit, ou ils peuvent être dispersés dans tout le réservoir. Si vous ne commencez pas à le combattre, alors avec l'aide de rhizoïdes, la culture s'accroche au substrat, comme si elle y poussait. Très souvent, ces algues apparaissent après l'achat de nouvelles plantes d'aquarium, ou si les règles d'entretien du bac sont négligées.
- Vietnamien. Ces algues d'aquarium sont des espèces filamenteuses. En fonction de leur apparence, les aquariophiles les appellent buisson, barbe ou pinceau. Les plantes viennent dans une variété de couleurs et se reproduisent très rapidement par les spores. La culture préfère s'asseoir sur les pointes des plantes d'aquarium ou décor de réservoir.
L'apparition de tout type d'algues indique des problèmes de microclimat dans le réservoir. Il faut des mois pour lutter contre certaines plantes, tandis que d'autres peuvent être éliminées rapidement et facilement.

3.2. Algues (Algues)
3.2.1. Les principales caractéristiques et la systématique des algues
Les algues sont un vaste groupe de plantes d'une grande importance biologique et très importantes pour l'humanité (section 3.2.8). Ce sont les plantes les plus primitives et n'ont pas de division corporelle en tige, racine et feuilles. Par conséquent, ils ont d'abord été combinés avec des champignons dans le département Thallophyta (voir note p. 43). Cependant, après de nouvelles découvertes scientifiques, il est devenu clair que les algues ne sont pas moins diversifiées que tous les autres groupes de plantes réunis, et qu'elles ont très peu caractéristiques communes. Il est probablement préférable de considérer les algues comme l'ensemble des organismes photosynthétiques producteurs d'oxygène ayant évolué dans le milieu aquatique et le maîtrisant parfaitement. Certes, certaines algues sont également venues se poser, mais à l'échelle planétaire, la productivité des formes côtières et terrestres est négligeable par rapport à la productivité des algues océaniques et d'eau douce. Si l'on adhère à ce point de vue, alors les algues bleues (Cyanophyta) devraient également être incluses dans le groupe des algues. Cependant, ces algues étant des procaryotes, il a été proposé de les appeler cyanobactéries (Cyanobactéries) afin de les distinguer en quelque sorte des algues eucaryotes. En même temps, un fait très important est négligé, à savoir que les algues bleu-vert libèrent de l'oxygène pendant la photosynthèse, contrairement à tous les autres procaryotes photosynthétiques. Pour que l'eau soit divisée en hydrogène et oxygène, la chlorophylle et le photosystème II sont nécessaires (section 9.4.2), ce qui constitue un avantage important par rapport aux bactéries photosynthétiques. On sait très peu de choses sur la façon dont cet avantage a été obtenu, bien que certaines formes intermédiaires entre les algues bleu-vert et les bactéries aient été trouvées. Cette interprétation de la relation entre les algues bleu-vert et d'autres plantes, y compris d'autres algues, est étayée par des preuves étayant la théorie symbiotique selon laquelle les chloroplastes végétaux proviennent d'algues bleu-vert (section 9.3.1).
En résumé, nous pouvons dire que le terme "algue" en lui-même est pratique, mais son utilisation en taxonomie introduit des complications inutiles. Les algues bleu-vert doivent être classées comme procaryotes et toutes les autres algues comme eucaryotes.
Heureusement, les algues eucaryotes appartiennent tout naturellement à des groupes bien définis, un ensemble de pigments photosynthétiques étant la principale caractéristique distinctive. Dans la systématique moderne, ces groupes ont reçu le statut de départements. Les relations entre départements ne sont pas encore élucidées, et cette question est très importante pour comprendre l'origine des plantes supérieures et la relation entre procaryotes et eucaryotes.
Tous les départements sont répertoriés dans la Fig. 3.11, et sur la fig. 3.12 donne des idées modernes sur les liens qui existent entre ces départements. Les principales caractéristiques des algues et certains des principaux départements sont indiqués dans le tableau. 3.4.

3.2.2. reproduction asexuée des algues
Les algues ont une reproduction asexuée et sexuée. Les principaux types de reproduction asexuée sont brièvement énumérés ci-dessous, du plus simple au plus complexe.
Multiplication végétative. Dans certaines formes coloniales, les colonies peuvent se diviser en fragments séparés, ce qui donne naissance à de nouvelles colonies plus petites. Dans les algues plus grandes, telles que Fucus, des thalles supplémentaires peuvent se former sur le thalle principal, qui se détachent et forment de nouveaux organismes.
Fragmentation. Ce phénomène est observé chez les algues filamenteuses telles que les algues bleues et Spirogyra. Le fil se divise d'une manière strictement définie et deux nouveaux fils sont formés. Ce phénomène peut être considéré comme l'une des formes de reproduction végétative.
Division binaire. Dans ce cas, un organisme unicellulaire se divise en deux moitiés identiques, tandis que le noyau se divise de manière mitotique. Une division longitudinale de ce type est observée à Euglena.
zoospores. Ce sont des spores mobiles avec des flagelles. Ils se forment dans de nombreuses algues, telles que Chlamydomonas, et dans certains champignons (voir Oomycota, tableau 3.2).
aplanospores. Ces spores immobiles se forment par exemple dans certaines algues brunes.
3.2.3. Reproduction sexuée chez les algues
La reproduction sexuée combine le matériel génétique de deux individus distincts de la même espèce. Le moyen le plus simple d'une telle reproduction est dans les algues; il consiste en la fusion de deux gamètes morphologiquement (c'est-à-dire structurellement) identiques. Un tel processus est appelé isogamie, et les gamètes isogamètes. Spirogyra isogame et certaines espèces de Chlamydomonas.
Si l'un des gamètes est moins mobile ou plus grand que l'autre, alors ce processus est appelé anisogamie. Chez Spirogyra, les gamètes ne diffèrent pas par leur structure, mais l'un d'eux bouge, tandis que l'autre est immobile. Cela peut être considéré comme une anisogamie physiologique. Il existe une autre option, lorsqu'un gamète est grand et immobile, et le second est petit et mobile. Ces gamètes sont appelés femelles et mâles, et le processus lui-même est appelé oogamie. Fucus oogamnas et certaines espèces de Chlamydomonas. Les gamètes femelles sont gros car ils contiennent un apport en nutriments nécessaires au développement du zygote après la fécondation.
Les trois types de reproduction sexuée correspondent à une augmentation de la complexité de la structure corporelle, et donc l'oogamie, bien que présente dans certaines algues simples, telles que Chlamydomonas, est généralement plus fréquente dans des algues plus complexes, telles que Phaeophyta. L'oogamie est le seul mode de reproduction sexuée chez les plantes qui sont plus hautement organisées que les algues.
Malheureusement, la terminologie utilisée pour décrire les gamètes et les organes reproducteurs chez les plantes est très confuse, en particulier chez les algues. Ci-dessous, nous n'expliquerons que les principaux termes.
Chez les champignons et les plantes inférieures (algues, bryophytes et fougères), les gamètes se forment dans des structures spéciales appelées gamétange. Le gamétange mâle s'appelle l'anthéridie et le gamétanium femelle s'appelle l'oogone ou archégone.
Oogonie* est un simple gamétange femelle présent dans de nombreuses algues et champignons, et les gamètes femelles ou les gamètes qui s'y trouvent sont appelés oosphères. L'oosphère fécondée est appelée oospore; il se transforme en une spore de repos à paroi épaisse, capable de survivre à des conditions défavorables. Nom commun pour le gamète femelle - Oeuf ou Oeuf, bien que parfois le terme « oosphère » soit utilisé pour désigner l'œuf ; cependant, ce n'est pas tout à fait exact.
* (Les oogones sont également appelées cellules ovariennes, à partir desquelles les ovocytes sont formés chez les animaux (voir chapitre 20).)
Archégone- il s'agit d'une gamétange féminine plus complexe, caractéristique des bryophytes, des fougères et de nombreux gymnospermes ; archegonium sera décrit plus loin dans ce chapitre.
À anthéridies des gamètes mâles sont produits, appelés anthérozoïdes ou spermatozoïdes. Ils sont mobiles car ils sont équipés d'un ou plusieurs flagelles. Ces gamètes sont caractéristiques des champignons, des algues, des bryophytes, des fougères et de certains gymnospermes. Chez les animaux, les gamètes mâles sont appelés spermatozoïdes ou sperme. Les noms listés sont montrés dans la fig. 3.13.

Pour les besoins de ce chapitre, il n'est pas si important de nommer les différents gamètes du même sexe, il suffit donc de faire la distinction entre les spermatozoïdes, c'est-à-dire tous les gamètes mâles, et les œufs, c'est-à-dire tous les gamètes femelles .
Comme pour les champignons, certaines algues présentent une hétérothallicité (section 3.1.3).
3.2.4. Division Chlorophyta
Les principales propriétés de Chlorophyta sont répertoriées dans le tableau. 3.4.

Tableau 3.4. Systématique et principales caractéristiques de quelques grands groupes d'algues 1)
1) (Un astérisque marque une caractéristique systématique.)
Chlamydomonas (chlamydomonas) est une algue mobile unicellulaire qui vit principalement dans les eaux stagnantes, c'est-à-dire dans les étangs et les fossés, surtout si l'eau est également enrichie en composés azotés solubles, comme les eaux de ruissellement des parcs à bestiaux. Les cellules de cette algue se trouvent souvent en si grand nombre que l'eau devient verte. Certaines espèces vivent dans l'eau de mer ou dans les estuaires saumâtres.
Structure
Chlamydomonas ne ressemble pas du tout à une plante, car elle bouge activement et a des vacuoles pulsantes. La structure de chlamydomonas est illustrée à la fig. 3.14. La micrographie électronique montre des organites eucaryotes typiques : l'appareil de Golgi, les mitochondries, les ribosomes et les petites vacuoles. Dans les chloroplastes de nombreuses algues, une structure spéciale a été révélée - pyrénoïde. Il s'agit d'une formation de protéines, constituée principalement de ribulose bisphosphate carboxylase, une enzyme qui fixe le dioxyde de carbone. Le pyrénoïde est impliqué dans le stockage des glucides, comme l'amidon. oeil rouge perçoit les changements d'intensité lumineuse, et la cellule se déplace là où l'intensité lumineuse est optimale pour la photosynthèse, ou reste en place si la lumière est suffisante. Cette réponse à la lumière s'appelle phototaxis(Chapitre 15.1.2). La cellule de chlamydomonas se déplace sous l'effet du battement de deux flagelles et est vissée dans l'eau comme un tire-bouchon, tournant autour de l'axe longitudinal.

Riz. 3.14. A. Chlamydomonas sous un microscope optique ; x 600. B. Schéma de la structure de Chlamydomonas. B. Micrographie électronique de Chamydomonas reinhardtii. × 1400
Cycle de la vie
Le cycle de vie de Chlomydomonas est représenté sur la fig. 3.15. L'adulte est haploïde.

reproduction asexuée
La reproduction asexuée est réalisée à l'aide de zoospores. La cellule mère perd des flagelles et le protoplaste de la cellule se divise en deux à quatre protoplastes filles (généralement quatre). En même temps, la division mitotique du noyau se produit; de plus, le chloroplaste se divise également. Les protoplastes filles développent de nouvelles parois cellulaires, de nouveaux yeux et de nouveaux flagelles. Les centrioles (corps basaux) sont impliqués dans la formation de nouveaux flagelles. La paroi cellulaire de la cellule mère devient mucilagineuse et les cellules filles, maintenant appelées zoospores, sortent. De chaque zoospore pousse une cellule de Chlamydomonas adulte à part entière. Ce processus est illustré à la Fig. 3.16, A

reproduction sexuée
Certaines espèces de Chlomydomonas sont homothalliques, d'autres sont hétérothalliques ; où différents types peut être isogame, anisogame ou oogame. La reproduction des espèces isogames est illustrée à la fig. 3.16, B. Pendant la germination, le noyau zygote se divise méiotiquement pour la première fois et l'état haploïde caractéristique des organismes adultes est restauré. Les jeunes cellules libérées de Chlomydomonas peuvent être appelées zoospores jusqu'à ce qu'elles soient complètement matures.
Dans les étangs et autres plans d'eau avec de l'eau stagnante mais claire, une autre algue vit - l'algue filamenteuse non ramifiée Spirogyra. La plupart des espèces de spirogyra sont des formes flottantes et ses fils sont visqueux et glissants.
Structure
Les cellules cylindriques de spirogyra sont connectées bout à bout et forment un fil illustré à la Fig. 3.17. Toutes les cellules sont identiques et il n'y a pas de séparation des fonctions entre elles. Une fine couche de cytoplasme se trouve le long de la périphérie de la cellule et une grande vacuole est, pour ainsi dire, enveloppée de brins de cytoplasme. Ces brins maintiennent le noyau au centre de la cellule. Un ou plusieurs chloroplastes en spirale se trouvent dans une couche à paroi mince du cytoplasme.

Croissance et reproduction
Les filaments de spirogyre se développent de manière intercalaire, c'est-à-dire en raison de la division de l'une des cellules qui composent le filament, quel que soit l'endroit où se trouve cette cellule. Chez la plupart des plantes, la zone de croissance est limitée à la région apicale. Le noyau de la cellule spirogyra se divise de manière mitotique, puis une nouvelle paroi cellulaire transversale est formée à partir des excroissances des parois latérales. Deux cellules filles sont obtenues, qui atteignent des tailles normales, à la suite de quoi le fil entier augmente en longueur.
Comme nous l'avons déjà noté (section 3.2.1), la reproduction asexuée se produit par fragmentation.
La reproduction sexuée s'effectue de manière très particulière, caractéristique des algues filamenteuses : deux filaments sont situés côte à côte et les cellules opposées des deux filaments sont reliées par de courtes excroissances tubulaires. L'ensemble du contenu de la cellule se comporte comme un gamète ; ce processus peut être considéré comme anisogame puisque, bien que les deux gamètes soient morphologiquement identiques, un seul d'entre eux est mobile et s'écoule dans l'autre cellule par un tube de liaison. Ce processus s'appelle la conjugaison.
3.2.5. Division Phaeophyta
Les principales caractéristiques de Phaeophyta sont répertoriées dans le tableau. 3.4.
Le long des côtes rocheuses de la côte britannique, on trouve souvent diverses algues du genre Fucus. Ils se sont très bien adaptés aux conditions assez rudes de la zone littorale, c'est-à-dire la zone qui est alternativement exposée à marée basse, puis à nouveau recouverte d'eau.
Les plus connues sont trois espèces de Fucus, que l'on trouve le plus souvent près de la côte dans trois zones différentes à des profondeurs différentes ; ce phénomène s'appelle distribution zonale. Ces algues sont classées en fonction de leur capacité à résister à l'exposition à l'air. Nous listons les principaux signes par lesquels ils peuvent être reconnus, et les endroits sur la côte où ils peuvent être trouvés :
F. spiralis (ces algues plates sont rejetées par la mer) - dans le point le plus haut marée haute. Lorsqu'il est immergé, le thalle est légèrement tordu en spirale.
F. serratus (ce qu'on appelle des algues ordinaires, en dents de scie ou en dents de scie) - dans la zone intertidale moyenne. Les bords du thalle sont dentelés.
F. vesiculosus (les soi-disant algues à bulles) - au point le plus élevé de la marée basse. Il y a des bulles d'air qui provoquent la flottabilité. Sur la fig. 3.18 vous pouvez voir la caractéristique signes extérieurs F. vesiculosus, et dans la fig. 3.19 montre les principales caractéristiques de sa structure interne.

Riz. 3.18. Structure externe de Fucus vesiculosus. Les traits caractéristiques et, en particulier, les adaptations à environnement. fin fructueuse(réceptacle) est une partie du thalle renflée et recouverte de petits renflements (scaphidies ou conceptacles), ne communiquant avec le milieu extérieur que par des trous étroits. Chez les plantes femelles, les extrémités fructifères sont vert foncé, chez les plantes mâles, orange. des bulles d'air généralement jumelés et donnent de la flottabilité aux algues. Branches adventices(parfois rompre; c'est une forme de multiplication végétative). cellule apex représente le point de croissance où se produit la division cellulaire. Bord- il s'agit d'une formation rigide qui remplit des fonctions mécaniques et, éventuellement, participe au transfert de certaines substances. Plaque plat et élastique (maigre); couleur brun verdâtre due à la couche photosynthétique proche de la surface ; recouvert de mucus, ce qui l'empêche de se dessécher à marée basse. La côte et la plaque forment un thalle. La partie d'enracinement du thalle (dans ce cas disque basal) est incolore et s'attache très fortement aux rochers avec un thalle, etc. La taille des algues varie jusqu'à 1 m ou plus. Thalle plat et en forme de ceinture; la nature de la ramification est telle que la résistance aux vagues est minimisée ; des bulles d'air soutiennent le thalle près de la surface, ce qui favorise la photosynthèse. Pétiole- c'est essentiellement une nervure; le pétiole est flexible et résiste donc avec succès aux vagues

Dans le corps de l'algue, ou thalle, il existe une certaine séparation des fonctions entre les différents tissus. Cette tendance est mieux observée chez Phaeophyta que dans tous les autres groupes d'algues. Nous verrons un peu plus loin les adaptations des algues à l'environnement.
Organes reproducteurs
La reproduction sexuée est oogame. F. vesiculosus et F. serratus sont dioïques, ce qui signifie qu'ils ont à la fois des mâles et des femelles. F. spiralis est un hermaphrodite, qui a des organes reproducteurs mâles et femelles sur la même plante dans les mêmes réceptacles - scaphidia ou conceptacles. Les organes reproducteurs se développent à l'intérieur des scaphidies sur les extrémités "fertiles" de certains thalles. Chaque scaphide a une ouverture étroite (pore) à travers laquelle les organes reproducteurs sont ensuite libérés. Leur structure est représentée sur la Fig. 3.19.
Les plantes adultes sont diploïdes et les gamètes se forment à la suite de la division méiotique.
Adaptations à l'environnement
Avant d'aborder les adaptations des Fucus à l'habitat, il convient de dire quelques mots sur le milieu lui-même, assez hostile. Étant des plantes de la zone intertidale, différentes algues sont exposées à l'air à marée basse à des degrés divers. Par conséquent, ils doivent disposer de dispositifs de protection contre le dessèchement. De plus, la température change très fortement lorsque les vagues froides de la mer se déversent dans les flaques chaudes laissées après la marée basse. Les plantes doivent s'adapter à un autre facteur, à savoir aux changements brusques de la salinité de l'eau, qu'elle soit augmentée par l'évaporation des petites mares formées après la marée basse, ou diminuée lors des pluies. Afin de résister à des facteurs tels que les marées, les vagues et les vagues, une résistance mécanique suffisante est nécessaire. De grosses vagues commencent à rouler des pierres, ce qui peut causer de très graves dommages aux plantes.
Adaptations morphologiques (structure générale)
L'algue thalle est solidement fixée au sol partie enracinée du thalle(rhizoïdes ou disque basal) (Fig. 3.18). Il se lie si fortement au sol (généralement des pierres) que les algues sont extrêmement difficiles à arracher. En règle générale, la pierre ne résiste pas à la première, et non à la partie d'enracinement du thalle.
Le thalle des algues n'est pas continu, mais disséqué; il se ramifie de manière dichotomique dans le même plan, ce qui vous permet de minimiser la résistance de la colonne d'eau. De plus, il est durable et résistant, mais pas rigide. Les côtes du thalle sont fortes et flexibles.
L'algue flottante F. vesiculosus possède des bulles d'air particulières qui maintiennent le thalle à la surface de l'eau, c'est-à-dire dans des conditions propices à une captation maximale de la lumière pour la photosynthèse.
adaptations physiologiques
Parmi les pigments photosynthétiques, le pigment brun prédomine - fucoxanthine. C'est l'une des adaptations pour la photosynthèse sous-marine, car la fucoxanthine absorbe fortement la lumière bleue, qui pénètre beaucoup plus loin dans la colonne d'eau que les longueurs d'onde plus longues, comme les rouges.
Le thalle sécrète beaucoup de mucus, qui remplit toutes les cavités internes de l'algue et suinte. Le mucus aide à mieux retenir l'eau et prévient la déshydratation.
La pression osmotique dans les cellules est beaucoup plus élevée que dans l'eau de mer, de sorte qu'aucune perte osmotique d'eau n'est observée.
Adaptations pour la reproduction sexuée
La libération des gamètes est synchronisée avec les marées. À marée basse, le thalle s'assèche et les organes reproducteurs sont expulsés des scaphidies, qui sont protégés du dessèchement par le mucus. Pendant la marée, les parois des organes reproducteurs se dissolvent, libérant des gamètes. Les gamètes mâles sont mobiles et ont un chimiotactisme positif pour les substances sécrétées par les gamètes femelles.
Le développement du zygote se produit immédiatement après la fécondation, ce qui minimise le risque d'être emporté dans l'océan.
3.2.6. Division Euglenophyta
Les principales caractéristiques d'Euglenophyta sont données dans le tableau. 3.4. Ce département est caractérisé par des signes de plantes et d'animaux, ce qui complique grandement la classification des organismes appartenant à cette zone. Pour cette raison, les botanistes et les zoologistes les incluent généralement dans leurs schémas systématiques. Ces problèmes seront discutés plus tard, après la description du genre Euglena.
Euglena est l'algue unicellulaire la plus commune trouvée dans les étangs d'eau douce, les fossés et tout autre plan d'eau riche en composés organiques dissous. Comme Chlamydomonas, elle se reproduit parfois si intensément que l'eau vire au vert, car la chlorophylle prédomine parmi les pigments de l'euglène. La structure d'euglena est illustrée à la fig. 3.20, où certaines de ses caractéristiques sont notées.

Riz. 3.20. Structure d'Euglena graciles. Canaliser- le lieu par lequel la nourriture entre dans les espèces non vertes ; la pellicule est absente ici, ce qui permet d'avaler de petites particules. judas(stigmatisation) est rouge ; impliqué dans la réaction de phototaxie. Photorécepteur détecte une source lumineuse et fait nager le corps dans la direction d'un éclairage optimal (phototaxis); la direction du mouvement peut changer lorsque le photorécepteur est ombragé. Flagelle long utilisé pour la locomotion; généralement dirigé vers l'avant ; des mouvements ondulatoires passent le long du flagelle de la base à la pointe; le flagelle entraîne la cellule derrière lui ; en avançant, la cellule tourne autour de son axe, laissant derrière elle une traînée en forme de tire-bouchon. Vacuole pulsatoire entouré de vacuoles accessoires ; participe à l'osmorégulation, pompant l'excès d'eau dans le réservoir, qui est entré dans la cellule à la suite de l'osmose. flagelle court ne participe pas à la locomotion. Granule de paramylone est formé par un polymère de glucose, similaire à l'amidon et est un glucide de stockage. pellicule situé sous membrane plasma; flexible. Chloroplastes contiennent des pigments photosynthétiques. À cytoplasme il y a des fibres contractiles qui provoquent des ondes péristaltiques de déformation cellulaire ; ce mouvement est appelé euglène
Euglena n'a pas de paroi cellulaire. A l'extérieur, la cellule est recouverte d'une membrane plasmique, juste en dessous de laquelle se trouve une protéine pellicule. La pellicule est assez flexible et cela permet à la cellule d'accepter forme différente. La pellicule entoure complètement le cytoplasme et peut être considérée comme une sorte de exosquelette. Il se compose d'une série de bandes longitudinales épaissies et de microfibrilles entrelacées. À l'intérieur du cytoplasme, de minuscules fibrilles appelées mionemy, les bandes de pellicule commencent à glisser les unes par rapport aux autres, ce qui modifie la forme du corps. Ce phénomène est appelé mouvement euglénoïde. Un autre moyen de locomotion, plus courant pour les euglènes, dû à la rotation d'un long flagelle, est illustré à la fig. 3.20 (considérez l'ocelle, le photorécepteur et le long flagelle) et est décrit en détail dans la sec. 17.6.3.
La reproduction asexuée se produit par la division longitudinale de la cellule en deux. La reproduction sexuée n'est pas observée.
Aliments
Les espèces vertes d'Euglena sont autotrophes et synthétisent toutes les substances dont elles ont besoin à partir du dioxyde de carbone, de l'eau et des sels minéraux. En même temps, ils ont besoin de recevoir des vitamines B1 et B12 de l'extérieur, qu'ils ne peuvent pas synthétiser eux-mêmes. En cela, Euglena n'est pas différente des animaux, bien que de nombreuses autres algues aient également un tel besoin de vitamines.
Plusieurs espèces d'Euglena n'ont pas de chlorophylle et ne sont donc ni colorées ni capables de photosynthèse (c'est-à-dire qu'elles sont hétérotrophes). Ils se nourrissent du type de saprophytes, la digestion se fait à l'extérieur de la cellule. Lorsqu'un plan d'eau est pollué, ils prospèrent, car le matériau en décomposition est riche en composés organiques. D'autres formes incolores sont capables d'avaler de petites particules de nourriture, pour lesquelles elles ont une sorte de "pharynx", où il n'y a pas de pellicule. Ces particules sont ensuite digérées à l'intérieur de la cellule (nutrition holozoïque, section 10.1.1). La nourriture est entraînée dans le pharynx par le mouvement des flagelles. Ces espèces rappellent à bien des égards les Rerapema les plus simples (Section 4.1.1).
Si les cellules vertes d'Euglena sont maintenues longtemps dans l'obscurité, les chloroplastes disparaissent et les cellules deviennent incolores. S'il y a suffisamment de substances organiques dans le milieu, les cellules peuvent vivre longtemps comme saprophytes. Lorsqu'elles sont mises au jour, la chlorophylle réapparaît.
Problèmes de taxonomie d'Euglena
Comme nous l'avons déjà dit, et comme il ressort du tableau. 3.1, Euglena se caractérise par des caractéristiques à la fois végétales et animales. L'un de ces signes animaux, auquel nous n'avons pas encore pensé, est la présence dans l'œil astaxanthine- un pigment caractéristique des animaux.
La facilité avec laquelle certaines euglènes peuvent passer du vert à l'incolore et vice versa indique que les espèces incolores en permanence semblent avoir évolué à partir du vert. Si par la suite les formes incolores ont développé des adaptations spéciales pour l'alimentation holozoïque, similaires à celles trouvées à Peranema, alors il est tout à fait possible que les ancêtres des protozoaires aient été similaires aux plantes. Il ne faut cependant pas oublier que l'évolution a pu aussi aller dans le sens inverse, puisque nous avons déjà évoqué au début de ce chapitre la possibilité que des ancêtres végétaux puissent être similaires à des animaux (c'est-à-dire des eucaryotes hétérotrophes).
Au moment de décider de placer Euglena dans le règne végétal ou dans le règne animal, il faut se rappeler que Chlamydomonas possède également certaines caractéristiques animales, et pourtant elle est généralement classée comme une plante. Les principales difficultés des taxonomistes sont liées à la méthode de nutrition. Apparemment, l'euglène devrait encore être attribuée aux plantes, car la présence de chloroplastes est considérée comme une caractéristique unique inhérente uniquement au règne végétal. Tout cela, cependant, nous rappelle une fois de plus combien il est difficile d'imposer à la nature une taxonomie artificielle inventée par l'homme.
3.3. Faites un tableau des personnages végétaux et animaux d'Euglena. Utilisez le tableau pour cela. 3.1, fig. 3.20 et les informations ci-dessus.
3.2.7. Directions d'évolution des algues
Même les quelques exemples que nous avons examinés dans les sections précédentes suffisent à comprendre qu'il existe de nombreux types d'algues, y compris des formes unicellulaires telles que Chlamydomonas, et des organismes relativement grands tels que Fucus, dans lesquels le corps est différencié et il existe une certaine fonctions de division entre les tissus individuels. Certaines grandes algues brunes ont même des tissus conducteurs, bien qu'elles n'aient pas de véritables tissus conducteurs - xylème et phloème.
Chez les algues, il existe une nette tendance à compliquer le processus de reproduction sexuée de la simple isogamie et anisogamie à l'oogamie. Cependant, l'une ou l'autre tendance doit être utilisée avec beaucoup de prudence pour expliquer les relations évolutives entre les différents groupes d'algues. De telles relations ne sont pas encore complètement élucidées et le groupe des Chlorophyta (algues vertes), dont seraient issues les plantes terrestres, est très diversifié : il présente à la fois des formes unicellulaires simples et des formes beaucoup plus complexes, et la reproduction sexuée varie également. de l'isogamie à l'oogamie.
3.2.8. La valeur des algues
Le rôle des algues dans la biosphère
Les estimations actuelles suggèrent que l'océan représente au moins la moitié de la production primaire mondiale en termes de carbone fixe. Cette production primaire est formée par les algues - les seules plantes qui habitent l'océan. Compte tenu de la vaste superficie occupée par l'océan, il faut s'attendre à ce que sa productivité soit encore plus grande, mais il ne faut pas oublier que la photosynthèse n'est possible que dans les couches superficielles, là où la lumière pénètre et où le facteur limitant est la disponibilité des nutriments, en particulier azote et phosphore.
Les algues sont des producteurs primaires très importants (Ch. 12) qui démarrent la plupart des chaînes alimentaires, y compris pratiquement toutes les chaînes marines et de nombreuses chaînes d'eau douce. Ces chaînes atteignent les poissons par le zooplancton*, les crustacés, etc. De nombreuses algues microscopiques sont unicellulaires et constituent le principal composant du phytoplancton * .
* (Le plancton est constitué des plus petits végétaux (phytoplancton) et animaux (zooplancton) qui nagent librement dans les couches superficielles des océans et des lacs. Le plancton a une grande importance économique et écologique.)
La fixation du carbone n'est qu'une des conséquences de la photosynthèse (section 9.2). De plus, la photosynthèse maintient le niveau d'oxygène dans l'atmosphère, avec au moins la moitié de tout l'oxygène produit par les algues, et leur contribution à ce processus est bien supérieure à celle des forêts terrestres.
Acide alginique, agar et carraghénane
De nombreux produits utiles sont obtenus à partir d'algues, tels que l'acide alginique, l'agar et le carraghénane. Acide alginique et ses dérivés (alginates) sont des polysaccharides extraits de la lame médiane et des parois cellulaires d'algues brunes telles que Laminaria, Ascophyllum et Macrocystis. Les algues sont récoltées en grande quantité dans les eaux côtières peu profondes ; Macrocystis, par exemple, est récolté au large de la Californie. Les alginates purifiés sont non toxiques et forment facilement des gels. Ils sont largement utilisés comme durcisseurs et gélifiants pour les produits industriels (par exemple, dans les cosmétiques - pour la fabrication de crèmes pour les mains) ; comme émulsifiants - pour la préparation de glaces; comme substances gélifiantes - dans l'industrie de la confiserie; dans la fabrication de vernis, peintures et médicaments ; pour obtenir des plats en céramique émaillée.
gélose- un polysaccharide obtenu à partir d'algues rouges. Il forme les mêmes gels que les alginates, mais est peut-être mieux connu comme un milieu très pratique pour la culture de bactéries et de champignons. À cette fin, une solution de gélose diluée est préparée, puis divers nutriments y sont ajoutés, stérilisés et laissés se solidifier, obtenant une masse gélatineuse. De plus, l'agar est utilisé aux mêmes fins que les alginates.
Carraghénane (carraghénane) est un autre polysaccharide de la paroi cellulaire dérivé principalement de l'algue rouge Chondrus crispus. Dans sa structure chimique, il est très similaire à l'agar et est utilisé aux mêmes fins.
Terre de diatomées (kieselguhr)
Les algues appartenant au département Bacillariophyta sont majoritairement unicellulaires ; elles sont appelées diatomées. Ces algues se caractérisent par une structure particulière de la paroi cellulaire, qui contient du silicium. Après la mort cellulaire, les restes de diatomées tombent au fond des mers et des lacs, et progressivement de gros dépôts s'y accumulent. La "terre de diatomées" ainsi formée contient beaucoup (jusqu'à 90%) de silicium. Après une purification appropriée, cette "terre" peut être utilisée comme excellent matériau filtrant (par exemple, dans la production de sucre ou pour clarifier la bière), comme charge dans la fabrication de peintures ou de papier, et comme matériau isolant capable de résister changements brusques de température.
Engrais
Dans les fermes situées près de la côte, les grosses algues (rouges et brunes) sont traditionnellement utilisées comme engrais, bien qu'à petite échelle. Les algues sont riches en potassium, mais elles contiennent beaucoup moins d'azote et de phosphore que dans le simple fumier. Par conséquent, leur effet fertilisant n'est pas très important. Un rôle plus important est joué par les algues bleu-vert libres, qui sont des fixateurs d'azote très importants et sont assez courantes dans le sol (section 9.11.1).
produits alimentaires
Certaines algues sont servies directement à table, notamment sur Extrême Orient. Considérées comme un mets délicat, l'algue rouge Porphyra et la grande algue brune Laminaria sont couramment consommées crues ou utilisées dans divers plats. Dans le sud du Pays de Galles, la porphyra est utilisée dans un plat traditionnel dans lequel des algues bouillies sont mélangées à de la farine d'avoine puis mijotées dans de l'huile. Dans la recherche de nouvelles sources de nourriture, une grande attention a été accordée à la culture industrielle des algues. Cependant, très peu d'algues se prêtent à l'obtention de nouveaux produits alimentaires, et jusqu'à présent aucun progrès significatif dans ce domaine n'a été réalisé dans la culture de bactéries et de champignons. Parmi les algues bleu-vert, la spiruline est considérée comme prometteuse.
Nettoyage des canalisations
Les algues apportent une certaine contribution au travail des micro-organismes dans le traitement des eaux usées, car les eaux usées contiennent des nutriments non seulement pour les bactéries, les champignons et les protozoaires, mais également pour les algues vertes microscopiques. Ils sont particulièrement utiles dans les "bassins d'oxydation" ouverts, qui sont largement utilisés dans les pays tropicaux et subtropicaux. Les bassins ouverts d'une profondeur de 1 à 1,5 m sont remplis d'eaux usées non traitées. Au cours de la photosynthèse, les algues libèrent de l'oxygène et assurent ainsi l'activité vitale d'autres micro-organismes aérobies se développant dans les eaux usées. De temps à autre, des algues sont récoltées et transformées pour l'alimentation du bétail.
Recherche scientifique
Les algues unicellulaires ont toutes les caractéristiques des plantes typiques, elles constituent donc un matériau idéal pour la recherche scientifique, car, d'une part, elles peuvent être cultivées en grand nombre dans des conditions strictement définies et, d'autre part, cela ne nécessite pas beaucoup d'espace. Un exemple de ces algues est Chlorella, qui occupe à juste titre une place d'honneur dans la recherche sur la photosynthèse (section 9.4.3). Les algues sont également utilisées dans l'étude de l'absorption des ions. Ils ont également été d'une grande utilité dans les études pionnières de la structure de la paroi cellulaire et des flagelles.
Les dégâts causés par les algues
Dans certaines conditions, les algues "fleurissent", c'est-à-dire qu'elles s'accumulent en grande quantité dans l'eau. La "floraison" est observée par temps assez chaud, lorsqu'il y a beaucoup de nutriments dans l'eau. Une telle situation est très souvent créée artificiellement par l'homme lorsque des effluents industriels sont déversés dans l'eau ou lorsque des engrais provenant des champs pénètrent dans les rivières et les lacs. En conséquence, la reproduction explosive des producteurs primaires (algues) commence et, en violation de toutes les lois de la nature, ils commencent à mourir avant d'avoir le temps d'être mangés. Avec la décomposition ultérieure des résidus, une reproduction tout aussi intensive de bactéries aérobies se produit et l'eau est complètement privée d'oxygène. Tout cela se produit très rapidement et, à cause du manque d'oxygène, les poissons et autres animaux et plantes commencent à mourir. L'augmentation de la concentration de nutriments dans l'eau qui déclenche tout ce processus s'appelle eutrophisation réservoir, et si cela se produit rapidement, nous pouvons supposer qu'il s'agit d'une autre forme de pollution de l'environnement.
Les toxines formées lors de la "floraison" de l'eau, notamment lors de la reproduction des algues bleues, augmentent la mortalité des animaux. Ces explosions d'algues sont un problème sérieux pour les fermes piscicoles, en particulier là où l'élimination intensive des engrais dans les champs augmente encore l'eutrophisation. Des complications similaires surviennent avec la "floraison" de l'eau dans l'océan. De plus, les toxines, s'accumulant dans le corps des mollusques et des crustacés qui se nourrissent d'algues, puis pénétrant dans le corps humain, provoquent divers empoisonnements et paralysies en lui.
Les algues sont également associées à de nombreuses difficultés pour stocker l'eau potable dans les réservoirs de réserve lorsqu'elle est contaminée par des déchets d'algues ou lorsque les algues commencent à se développer sur les filtres à sable, les obstruant complètement.
3.4. Les difficultés dont nous venons de parler sont plus susceptibles de se produire dans les réservoirs situés dans les basses terres. Expliquez pourquoi c'est ainsi que cela devrait être.
3.5. Contrairement à de nombreux champignons et bactéries, les algues ne causent aucune maladie. A quoi est-ce lié ?
Le manuel est rédigé conformément aux exigences de la norme fédérale d'enseignement de l'enseignement professionnel supérieur dans le sens de "l'éducation pédagogique" et complète les connaissances des étudiants dans la partie théorique du cours "Botanique" (systématique des plantes et des champignons) . Le matériel du manuel peut être utilisé par les étudiants à la fois pour un travail indépendant et pour un travail en classe sous la direction d'un enseignant.
* * *
L'extrait suivant du livre Botanique. Plant Systematics: a textbook (S. K. Pyatunina, 2013) fourni par notre partenaire de livre - la société LitRes.
Algue (algues)
Un groupe important et diversifié de plantes à thalles inférieurs dont l'habitat principal est l'eau. Algae réunit plusieurs départements indépendants et, selon toute vraisemblance, évolués indépendamment. Les représentants des départements diffèrent par l'ensemble des pigments, les détails de la structure fine des chromatophores, les produits de la photosynthèse qui s'accumulent dans la cellule (substances de réserve) et la structure de l'appareil flagellaire. Les plantes inférieures sont des organismes unicellulaires, coloniaux ou cénobiens et multicellulaires. Les colonies sont appelées cénobies, dans lesquelles le nombre de cellules est déterminé dans les premiers stades de développement et ne change pas avant la prochaine étape de reproduction (reproduction). La croissance des coenobia se produit en raison d'une augmentation de la taille des cellules, et non de leur nombre. Il existe les types suivants d'organisation morphologique du thalle:
1. monadique- des cellules qui se déplacent activement à l'aide de flagelles.
2. coccoïde- cellules immobiles.
3. rhizopodial (amiboïde)- les cellules végétatives ne sont pas recouvertes de membranes et peuvent développer des processus cytoplasmiques - rhizopodes.
4. palmelloide, ou capsale, le type d'organisation est représenté par des cellules immobiles immergées dans un mucus commun.
5. filamenteux- des cellules reliées en fils, simples ou ramifiées.
6. hétérotriches ou multifilamenteux,- une version compliquée de la structure filamenteuse, caractérisée par deux systèmes de fils: rampant le long du substrat et fils verticaux s'étendant à partir de ceux-ci.
7. lamellaire- des thalles en forme de plaques.
8. Siphonal- les thalles, souvent de grande taille, représentent formellement une cellule, généralement avec un grand nombre de noyaux.
9. siphonocladale l'organisation est représentée par des cellules multinucléaires reliées par des filamenteux ou d'autres formes de thalles multicellulaires. Aux premiers stades de la formation du thalle, il a une structure de type siphonal.
Les algues peuvent se reproduire de trois manières : végétative, asexuée et sexuée. La multiplication végétative consiste à séparer une partie du thalle végétatif de la plante entière, donnant naissance à un nouveau thalle. La reproduction asexuée est réalisée à l'aide de cellules spécialisées - contestation, produit dans les sporanges. Les spores sont mobiles (zoospores) ou immobile (aplanospores). Ils peuvent être de forme identique aux thalles parents. (autospores algues unicellulaires) ou s'en distinguent fortement (spores unicellulaires d'algues multicellulaires).
La reproduction sexuée chez les algues est extrêmement diversifiée. Les formes les plus simples du processus sexuel - la fusion d'individus végétatifs morphologiquement indiscernables - hologamie et conjugaison. Dans une partie importante des algues, la formation de cellules germinales spécialisées - gamètes. On distingue le comportement suivant des hemets :
1. Isogamie - fusion de gamètes de même taille et de même forme.
2. Hétérogamie - les deux types de gamètes en copulation ont des flagelles, mais la femelle est plus grande et moins mobile que le mâle.
3. Oogamie - fusion d'un œuf femelle immobile et d'une cellule mâle mobile. Copuler les gamètes qui sont apparus sur le même individu (homothallisme) ou sur des individus différents (hétérotallisme). L'hétérotallisme est observé dans toutes les formes du processus sexuel. Dans les formes isogames, les gamètes à identité morphologique se révèlent physiologiquement différents et sont désignés par les signes conventionnels "+" et "-". Les gamètes mâles porteurs de flagelles sont appelés spermatozoïdes, ne pas avoir de flagelles, mais capables de se déplacer à l'aide de mouvements amiboïdes sont appelés spermatozoïdes.À la suite du processus sexuel, une cellule diploïde se forme - zygote.
Cycle de la vie, ou cycle de reproduction des algues, comprend la croissance végétative, la sporulation asexuée, le processus sexuel, les stades de repos. Le rapport des phases diploïdes et haploïdes dans le cycle de vie des algues n'est pas le même. Dans certains cas, la germination du zygote s'accompagne d'une division de réduction (méiose) du zygote. (réduction zygotique), tandis que les plantes en développement sont haploïdes. Chez de nombreuses algues vertes, le zygote est le seul stade diploïde du cycle de développement ; toute la phase végétative passe à l'état haploïde. Ce cycle de vie est appelé monohaplobionte. Chez certaines autres algues, au contraire, toute la phase végétative est diploïde, la phase haploïde n'est représentée que par des gamètes, avant la formation desquels se produit la division par réduction du noyau. (réduction gamétique), cycle de la vie - monodiplobiontique. Dans d'autres encore, la division de réduction du noyau précède la formation de spores qui se développent sur les thalles diploïdes. (réduction sporique). Ils se transforment en plantes haploïdes à reproduction sexuée. (gamétophytes). Après la fusion des gamètes, le zygote se développe en une plante diploïde, portant des organes de reproduction asexuée ( sporophytes). Ainsi, ces algues ont une alternance de générations (générations) : diploïdes et haploïdes. Cycle de la vie - haploïde-diploïde. Les deux générations peuvent être morphologiquement identiques ( changement isomorphe de générations) ou d'apparence radicalement différente (changement hétéromorphe de générations).
En cours pratiques, ils étudient les départements : algues vertes (Chlorophyta), diatomées (Bacillariophyta ou Diatomeae), brunes (Phaeophyta), rouges (Rhodophyta).
Département des algues vertes (Chlorophyta)
Le département des algues vertes est le plus grand en termes de nombre d'espèces (jusqu'à 20 000 espèces) et de département d'algues morphologiquement diversifié. Il existe également de petites formes unicellulaires microscopiques (monadiques et coccoïdes) et des formes filamenteuses, hétérotriches, siphonales, siphonocladales et lamellaires assez complexes, atteignant plusieurs dizaines de centimètres. L'aire de répartition des algues vertes est également étendue (on les trouve partout dans le monde) et leur amplitude écologique est large. Ils vivent dans les eaux douces et marines, certains vivent hors de l'eau. Mais avec toute la diversité, les algues vertes ont un certain nombre de caractéristiques communes :
1) composition pigmentaire : chlorophylle un et dans, les caroténoïdes et les xanthophylles ;
2) le principal produit de réserve de nature glucidique, l'amidon, se dépose dans le chromatophore autour du pyrénoïde ;
3) œil photosensible - stigmate, situé dans le stroma du chromatophore;
4) les thylakoïdes portant des pigments ont tendance à s'empiler ;
5) les flagelles sont isomorphes (de même structure) et isocontoïdes (de même longueur).
Classe En fait vert, ou flagelles égaux, algues (Chlorophycées, Isocantae)
Les représentants de cette classe se caractérisent par une reproduction asexuée à l'aide d'aplanospores immobiles ou de zoospores mobiles avec deux à quatre, moins souvent de nombreux flagelles isocont et isomorphes. Processus sexuels - chologamie ou copulation de gamètes - isogamie, hétérogamie, oogamie. Le zygote passe généralement par un état dormant et germe lorsque des conditions favorables se présentent, et son noyau diploïde se divise immédiatement par réduction. Conformément aux étapes de différenciation morphologique du thalle, la classe est divisée en ordres.
Commander Volvox (Volvacales)
L'ordre comprend des algues unicellulaires, coloniales et cénobiennes équipées de flagelles, c'est-à-dire une organisation monadique.
Genre Chlamydomonas (Chlamydomonas)
Chlamydomonas est un genre étendu d'environ 500 espèces, largement distribué dans la nature. Ses espèces peuvent être trouvées dans des réservoirs peu profonds et bien chauffés, des flaques d'eau, des fossés. Avec un développement massif, il provoque la floraison de l'eau, en particulier dans les réservoirs pollués par des substances organiques. Le thalle des chlamydomonas est unicellulaire, d'organisation monadique, c'est-à-dire qu'étant dans un état actif, les chlamydomonas se déplacent rapidement à l'aide de deux flagelles égaux attachés à l'extrémité antérieure du corps. La phase de mouvement actif est remplacée par un état de repos. C'est le stade dit palmelle-like, lorsque les cellules perdent leurs flagelles, leurs membranes deviennent fortement mucilagineuses et forment des agrégations de cellules de Chlamydomonas immergées dans le mucus commun. Sous cette forme, les cellules de chlamydomonas se multiplient par division. Entrant dans des conditions d'existence favorables, les chlamydomonas produisent à nouveau des flagelles et passent à un mouvement actif.
Chlamydomonas a une membrane cellulaire de cellulose-pectine, un chromatophore en forme de coupe avec un ou plusieurs pyrénoïdes situés dans la partie inférieure et un œil sensible à la lumière (stigmate) dans la partie supérieure. Le noyau est situé dans l'approfondissement du chromatophore, il y a une paire de vacuoles pulsantes. La reproduction asexuée par les zoospores se produit dans des conditions d'habitat favorables. Chaque Chlamydomonas peut potentiellement se reproduire de manière végétative et asexuée, ainsi que participer au processus sexuel. Lors de la reproduction asexuée, le protoplaste se divise en 4 ou 8 parties, des zoospores se forment. Le processus sexuel chez la plupart des espèces est isogame. Les gamètes se forment de la même manière que les zoospores, mais en plus grand nombre (32 ou 64). Le zygote est bien adapté pour supporter des conditions défavorables. Sa germination s'accompagne d'une division de réduction. Le cycle de développement de chlamydomonas est monohaplobiont.
Rod Volvox (Volvox)
Le genre Volvox est une algue coloniale ou cénobienne. Un petit genre de Volvox vit dans les eaux stagnantes propres, les étangs et les petits lacs. C'est le représentant le plus organisé de l'ordre Volvox. C'est une grosse boule atteignant 2 à 3 mm de diamètre, recouverte d'une fine couche de mucus (involucrum), sous laquelle des cellules biflagellées sont situées dans une couche le long de la périphérie de la boule. Leur nombre varie de 500 à 60 000. La cavité interne de la balle est occupée par du mucus liquide. Les cellules de la colonie ont une structure similaire aux cellules de chlamydomonas. Mais la coquille de chaque cellule est fortement mucus, de sorte que les protoplastes des cellules voisines sont éloignés les uns des autres et les processus cytoplasmiques pénètrent dans l'épaisseur de la membrane muqueuse. Des plasmodesmes se forment aux points de contact.
Dans la reproduction asexuée, 8 à 10 cellules sont impliquées, situées à l'arrière, par rapport à la direction du mouvement, une partie de la sphère. Ce sont des gonidies. Entre autres, ces cellules se distinguent par des tailles plus grandes. Lorsqu'ils se divisent, une plaque plate de 16 cellules (stade gonique) est d'abord formée, une division supplémentaire conduit à la formation d'une sphère ouverte avec un petit trou ouvert dirigé vers la surface externe de la colonie mère. Les cellules en formation du nouvel organisme sont tournées avec leurs flagelles dans la sphère. L'orientation normale des cellules (avec les extrémités antérieures dirigées vers l'extérieur) est obtenue en tournant complètement la sphère ouverte à l'envers, seulement après que son ouverture se ferme. Les cellules reproductrices se différencient très tôt, de sorte que non seulement des colonies filles, mais aussi des colonies petites-filles peuvent être observées à l'intérieur de l'organisme maternel. Les jeunes colonies sont libérées après la destruction de la colonie mère.
Les cellules qui servent à la reproduction sexuée sont les oogones et les anthéridies. Les ovogonies vert foncé sont beaucoup plus grandes que les autres cellules et manquent de flagelles. Un gros œuf se développe dans l'oogone. Les anthéridies forment des paquets de spermatozoïdes. Le processus sexuel chez Volvox est oogame. Il existe des espèces bisexuées et dioïques, ainsi que des clones homo et hétérothalliques. Un zygote au repos est formé, qui germe en tant que jeune colonie fille après la division par réduction des noyaux diploïdes. Le cycle de développement est monohaplobionte.
Objets : r. chlamydomonas, r. volvox.
Progrès
1. Considérez d'abord les chlamydomonas à faible grossissement (m. grossi), puis plus en détail à fort grossissement (b. grossi), étudiez les individus immobiles, observez le mouvement des chlamydomonas.
2. Faites deux dessins :
a) l'apparition de chlamydomonas. Désigner la coquille, chromatophore ;
b) un schéma de la structure de la cellule de chlamydomonas, à l'aide du tableau. Désigner la membrane, le cytoplasme, le noyau, le chromatophore, l'ocelle (stigmate), le pyrénoïde, les flagelles, les vacuoles pulsatoires.
3. Considérez en m. et esquissez des coenobia sphériques de volvox avec des colonies filles de la préparation.
4. Faites un dessin schématique à l'aide du tableau, reflétant les caractéristiques structurelles du coenobia. Nommez les protoplastes, les processus cytoplasmiques, les plasmodesmes, les flagelles, les ovogonies, les anthéridies.
Commander Chlorocoque (Chlorococcie)
L'ordre comprend des formes unicellulaires et cénobiennes avec une organisation cellulaire coccoïde.
Genre Chlorococcus (Chlorocoque)
Le genre Chlorococcus contient 38 espèces et se trouve dans une variété d'habitats : dans l'eau, à la fois dans le plancton et dans le benthos ; dans le sol, ainsi que sur l'écorce des arbres, sur les vieux bâtiments en bois. Le chlorocoque fait partie des lichens.
C'est une algue coccoïde unicellulaire, immobile à l'état végétatif. Les cellules ont un chromatophore en coupe avec un pyrénoïde, mais manquent de flagelles, d'ocelles et de vacuoles pulsantes. Le noyau est situé dans la cavité du chromatophore. Chez les individus âgés, plusieurs noyaux peuvent être observés, les cellules sont recouvertes d'une épaisse membrane de cellulose.
Chlorococcus se reproduit de manière asexuée à l'aide de zoospores biflagellées allongées. Le protoplaste de la cellule mère se divise et forme de 8 à 32 zoospores, qui sont libérées après la destruction de la paroi de la cellule mère. La période de mouvement actif est courte; après avoir nagé pendant un certain temps, les zoospores perdent leurs flagelles, s'habillent d'une coquille, grandissent et atteignent des tailles caractéristiques d'une espèce particulière. Le processus sexuel est isogame. Le cycle de développement est monohaplobionte.
Genre Hydrodition (Hydrodiction)
Hydrodition est un genre petit mais répandu. On le trouve dans les eaux stagnantes des rivières, des étangs et autres plans d'eau stagnante enrichis en azote. Il s'agit d'une algue cénobienne macroscopique, composée d'un grand nombre (jusqu'à 20 000) de cellules. Les vieux spécimens atteignent un mètre de long et leurs cellules mesurent jusqu'à un centimètre et demi.
L'hydrodiction ressemble à un réseau fermé, composé de 5-6 cellules de charbon formées par des cellules géantes reliées à leurs extrémités. Les cellules du réticulum adulte contiennent une vacuole géante, le cytoplasme est pariétal et contient un chromatophore réticulé avec de nombreux pyrénoïdes et un grand nombre de petits noyaux. La coque est en cellulose. Chaque cellule assure toutes les fonctions de l'organisme (nutrition et reproduction).
La reproduction de la maille d'eau est asexuée et sexuée. Les cellules qui ont déjà atteint une taille suffisamment grande (0,2 mm) commencent la reproduction asexuée. Dans le protoplaste, un tel nombre de zoospores est formé, caractéristique de ce type de réticulum. Les zoospores ne quittent pas la cellule mère, mais se déplacent à l'intérieur de la cellule pendant un certain temps.
Ensuite, les flagelles sont jetés et pliés en un jeune maillage, collés les uns aux autres aux endroits où passent les brins de microtubules. Les cellules du jeune réticulum sont mononucléaires, avec un chromatophore lamellaire portant un pyrénoïde. Pendant un certain temps, le jeune maillage vit sous la coquille de la cellule mère, mais sa taille augmente rapidement, les cellules s'allongent. En fin de compte, la coquille de la cellule mère est détruite et le réticulum commence à vivre de manière indépendante.
La reproduction sexuée est isogame, les gamètes sont plus formés que les zoospores et ils sont beaucoup plus petits. Le zygote est coloré avec du rouge brique hématochrome. Après une période de dormance, le zygote se divise par réduction et germe en quatre grandes zoospores. Ils sont inactifs, perdent bientôt leurs flagelles et se recouvrent à nouveau d'une coquille épaisse, mais déjà sculpturale, se transformant en soi-disant polyèdre Les polyèdres, dont les coquilles ont des processus et dont le contenu est riche en inclusions grasses, sont apparemment importants pour la distribution des algues. Les polyèdres supportent bien la dessiccation et sont donc la deuxième étape dormante du cycle de développement du réseau d'eau.
Genre Chlorelle (Chlorelle)
La chlorelle est une algue très répandue. Dans la nature, on le trouve dans le plancton et le benthos de divers plans d'eau, sur le sol, participe à la formation du corps des lichens et vit également en symbiose avec de petits animaux, formant la soi-disant zoochlorelle. Elle fait partie des algues cultivées. En raison de son taux de reproduction élevé, la chlorella donne un rendement élevé en biomasse.
La chlorelle est une algue coccoïde sphérique unicellulaire. Le protoplaste contient un chromatophore en forme de cloche avec une grande dépression. Dans la cavité du chromatophore, un noyau peut être trouvé. Chlorella se reproduit uniquement de manière asexuée par les autospores. Le processus sexuel est inconnu. Le cycle de développement est asexué, monohaplobionte.
Objets : r. chlorocoque, r. maille d'eau, r. chlorelle.
Progrès
1. Considérez en b. emmené. et dessine le chlorocoque. Désigner une coquille épaisse (une adaptation à un mode de vie aérophile), un chromatophore en forme de coupe.
2. Considérez en m. un fragment du coenobium du maillage de l'eau et dessin, montrant des cellules à 5-6 angles qui forment le coenobium. Désigner la cellule de coenobia, chromatophores.
3. Considérez en b. emmené. et dessinez une cellule d'un coenobium avec un jeune coenobium à l'intérieur de la cellule. Désigner la coquille de la cellule mère, le jeune coenobium.
4. Esquissez la chlorella en b. emmené. Nommez la coquille épaisse, chromatophore.
Commander Ulotrix (Ulotrichales)
L'ordre regroupe des algues qui ont un thalle en forme de fil non ramifié, composé de cellules mononucléaires, moins souvent un thalle lamellaire ou tubulaire.
Canne Ulotrix (Ulothrix)
Ulothrix est un genre assez grand, trouvé dans les plans d'eau douce et légèrement saumâtre, préférant l'eau courante propre. Dans la zone côtière des rivières et des ruisseaux, en particulier dans les zones au climat approprié, on peut trouver Ulothrix zonata. Ulothrix est une algue benthique attachée qui forme des agrégations sur les roches côtières.
Ulothrix est une algue multicellulaire filamenteuse non ramifiée. Toutes les cellules, à l'exception de la basale, qui sert à l'attachement, sont du même type. Ils sont recouverts d'une fine membrane cellulosique parfois mucilagineuse. Le chromatophore est pariétal sous la forme d'une ceinture fermée ou ouverte avec un grand nombre de pyrénoïdes. Il n'y a qu'un seul noyau dans les cellules. Le nombre de cellules dans les filaments est indéterminé, car les cellules des parties supérieures se divisent constamment dans le même plan. Ulothrix se reproduit végétativement (par fragmentation du fil), asexuée et sexuée. La reproduction asexuée est réalisée par des zoospores équipées de quatre flagelles isocontes et isomorphes. Les zoospores, en germination, forment des filaments ulotrix. Toute cellule sauf la cellule basale du filament peut potentiellement devenir un zoosporange. Ulothrix forme des isogamètes biflagellés. Après la fusion, un planosygote est formé (une cellule mobile à quatre flagelles), qui perd alors les flagelles et se différencie en un sporophyte particulier unicellulaire - un corps élargi recouvert d'une coquille épaisse sur jambe mince. Le zygote-sporophyte reste dans un tel état de repos pendant un certain temps, puis, après division par réduction, 4 à 16 zoospores s'y forment. Un tel cycle de vie est caractéristique d'Ulothrix zonata. Chez certaines espèces, le zygote se transforme en un filament diploïde. Ainsi, le cycle de développement chez Ulothrix zonata est avec un changement hétéromorphe d'un sporogamétophyte haploïde filamenteux multicellulaire et d'un sporophyte diploïde unicellulaire.
Tige Ulva (Ulva)
Le genre Ulva est connu sous le nom de "laitue de mer" et est largement distribué mais préfère les eaux peu profondes. Ulva est une algue, mais elle tolère bien le dessalement. Il vit dans les estuaires, les estuaires peu profonds, les marais ; tolère bien et préfère en partie les eaux à forte pollution organique. La population locale l'utilise comme nourriture, mais l'ulve n'a aucune valeur commerciale.
Le thalle de l'ulve est multicellulaire, lamellaire, constitué de deux couches de cellules, les bords de la plaque sont ondulés en raison de divisions cellulaires plus intenses dans les zones marginales par rapport à la zone médiane. À la base, la plaque se rétrécit en un court pétiole avec une semelle, avec laquelle elle est attachée à un substrat solide. La différenciation cellulaire dans le thalle de l'ulve est faible. Il n'y a pas d'organes reproducteurs spéciaux. Potentiellement, chaque cellule peut devenir un sporange dans la génération diploïde ou un gamétange dans la génération haploïde. Certaines cellules ont des excroissances tubulaires descendant le long de la partie centrale du thalle.
La reproduction asexuée est réalisée par des zoospores à quatre flagellations, qui se forment sur des plantes diploïdes après division par réduction. Les zoospores germent en un fil non ramifié à une seule rangée, mais avant même le début de la division, une polarisation est détectée dans la zoospore qui a perdu des flagelles et s'est fixée au sol. L'extrémité supérieure est plus épaisse, tandis que l'extrémité inférieure est mince et allongée, à partir de laquelle des structures de fixation sont ensuite formées. Le processus sexuel est iso- ou hétérogame, les gamètes sont biflagellés. Le cycle de développement est haplodiplobiont avec un changement isomorphe de générations.
Objets : r. Ulotrix, r. ulve.
Progrès
1. Considérez en b. emmené. et dessinez une section du filament ulotrix, faites attention à la structure du chromophore. Étiqueter les cellules du thalle : membrane cellulaire, chromatophore, pyrénoïdes.
2. Dessinez l'apparence du thalle de l'ulve à l'aide d'une préparation humide.
Ordre des chaétophores (Chaetophorales)
L'ordre comprend des formes filamenteuses multicellulaires de type hétérotriche avec différenciation du thalle en un système horizontal, réparti sur le substrat, et un système vertical de filaments.
Rod Draparnaldia (Draparnaldia)
Les espèces de ce genre exigent la propreté et l'aération des réservoirs et préfèrent les rivières et les ruisseaux à débit rapide. Ils poussent en masse à des profondeurs assez importantes (10 m), y formant des fourrés entiers. Draparnaldia benthic est une algue attachée de structure hétérotricheuse filamenteuse. Il a de longs filaments (croissance illimitée) faiblement ramifiés avec un chromatophore en forme de ceinture avec des bords dentelés. Le chromatophore dans ces branches est petit par rapport au volume total de la cellule, de sorte que les cellules des filaments principaux sont pâles. Des bouquets de branches courtes très ramifiées à croissance limitée partent de ces filets en verticilles, ce sont des assimilateurs. Les chromatophores qu'ils contiennent sont pariétaux, grands; cellules vertes. Chaque fil court se termine par un poil long incolore. Les organes reproducteurs sont placés parmi les assimilateurs. Reproduction asexuée par quatre zoospores flagellaires. Le processus sexuel est l'isogamie ou l'hétérogamie. Le cycle de développement est monohaplobionte.
Genre Trentepolia (Trentepohlia)
Trentepolia est une algue terrestre aérophile, bien adaptée au manque d'humidité. Elle s'installe sur l'écorce des arbres, des pierres, des constructions en bois. Surtout de nombreuses espèces de ce genre se trouvent dans les régions tropicales et subtropicales humides, où trentepolia mène souvent un mode de vie épiphyte. Trentepolia est facile à reconnaître parmi les autres algues terrestres grâce à l'hématochrome, un caroténoïde soluble dans l'huile qui lui donne une couleur rouge brique ou jaune. Le thalle de trentepolia est filamenteux, hétérotriche. Les filaments qui rampent le long du substrat sont constitués de cellules arrondies ou ovales recouvertes d'une épaisse membrane en couches. Ils sont reliés les uns aux autres par des pores avec des plasmodesmes, mais, néanmoins, les filaments rampants se désintègrent facilement en fragments courts ou en cellules individuelles. Ces fragments et cellules à l'état sec sont dispersés et emportés par le vent. Ainsi, trentepolia a un mécanisme de reproduction végétative assez efficace, ce qui est particulièrement important dans des conditions de carence en eau. Les cellules contiennent plusieurs chromatophores en forme de disque ou de ruban dépourvus de pyrénoïdes et un grand nombre de noyaux, en particulier dans les cellules âgées. En plus de l'horizontal, il existe un système de fils verticaux, constitué de cellules plus allongées. Les filaments horizontaux et verticaux se ramifient abondamment en raison de la division des cellules apicales ; des coiffes en couches se forment sur ces dernières.
La reproduction asexuée se produit par des zoospores à deux ou quatre flagellations, qui se forment dans des cellules apicales spéciales - les sporanges, assises sur des cellules tubulaires - les jambes. Les sporanges sont détachés et emportés dans leur intégralité par les courants d'air. Les zoospores ne se forment que si le sporange est dans l'eau. Ensuite, le contenu multinucléaire très rapidement, en quelques minutes, se décompose en sections mononucléaires et des zoospores sont produites. Les gamétanges diffèrent également morphologiquement des cellules végétatives, mais sont principalement situées sur des filaments rampants. Contrairement aux sporanges, les gamétanges globulaires n'ont pas de pattes. Les gamétanges sont également transportés par les courants d'air. Une fois dans l'eau, ils germent avec des isogamètes biflagellés. Cependant, la copulation est rare et les gamètes se développent parthénogénétiquement (sans fécondation). Dans le cas de la formation de zygotes, ils germent sous forme de zoospores après une période de dormance. Le cycle de développement est monohaplobionte.
Objets : r. draparnaldia, r. trentepoly.
Progrès
1. Considérez en m. et dessinez une section du thalom filamenteux hétérotrichal de Draparnaldia, notez les caractéristiques structurelles des filaments et des assimilateurs de la «tige», montrez la différence de structure des chromatophores dans les cellules du filament axial et dans les cellules des branches latérales. Désigner le filetage axial, les filetages assimilateurs.
2. Considérez en b. emmené. et tracer la cellule du filetage axial et la cellule du filetage assimilateur. Étiquetez la paroi cellulaire, le chromatophore.
3. Considérez en b. emmené. et esquissez une section du fil trentepoly. Désigner la membrane en couches, les substances de réserve (sous forme de gouttes d'huile colorées à l'hématochrome), les chromatophores en forme de disque.
Ordre des Cladophoracées (Cladophorales)
L'ordre comprend des algues non cellulaires, ramifiées filamenteuses, divisées par des cloisons transversales en segments inégaux, dont chacun contient de nombreux noyaux. Les cloisons transversales apparaissent indépendamment de la fission nucléaire.
Genre Cladophore (Cladophore)
Cladophora est un genre très vaste et répandu d'algues principalement marines et en partie d'eau douce. Environ 150 espèces ont été décrites. Il se produit dans les eaux peu profondes de la zone de surf, sur les rochers faisant saillie dans la mer, dans les lagons, les étangs, les lacs. Les jeunes plantes sont attachées au sol ou à divers objets sous-marins, mais se détachent plus tard et nagent, formant de grandes grappes ou de la boue dure; ainsi que de grands amas globulaires (10–15 cm de diamètre). Le thalle est très ramifié, avec une structure recouverte de siphon. Les segments formés par les cloisons transversales ne sont pas des cellules au sens propre. La formation de septa transversaux est qualitativement différente de la cytokinèse et n'est pas associée à la fission nucléaire. Les segments sont de taille inégale et contiennent un nombre différent de cœurs. Le chromatophore maillé est également formé par contact et fusion de chloroplastes initialement libres. L'enveloppe extérieure est épaisse, en cellulose, jamais de mucus, donc la boue de Cladophora est dure et non glissante.
La reproduction est asexuée et sexuée. N'importe quel segment de l'algue peut devenir zoosporange ou gamétange. Les zoospores tétraflagellées émergent du sporange par un pore et germent d'abord dans un corps vésiculaire dépourvu de septa (stade siphonal), puis des septa transversaux apparaissent et une ramification des structures filamenteuses se produit. Le processus sexuel est isogame. Le zygote devient une plante diploïde. La plupart des espèces de cladophores vivant dans les mers ont un changement isomorphe de générations, auquel cas les zoospores se forment après la division par réduction (méiozoospores), mais les espèces d'eau douce ont un cycle monodiplobionte, lorsque la division par réduction précède la formation des gamètes.
Objets : r. cladophore.
Progrès
Considérons en b. emmené. et dessinez une section du thalle ramifié du cladophore avec des zoosporanges. Désigner les cellules du thalle, les zoosporanges unicellulaires.
Conjugués de classe (Conjugatophycées)
Les représentants de cette classe ont un type particulier de processus sexuel - conjugaison, il n'y a pas de stades flagellaires, il n'y a pas de reproduction asexuée par les spores.
Commande Zignemov (Zygnematales)
Commandez Zignemovye - algues multicellulaires filamenteuses non ramifiées, leur coquille est mucilagineuse et elles sont donc glissantes au toucher. Le zygote germe sous la forme d'un semis, les trois autres noyaux formés lors du processus de division par réduction meurent.
Genre Spirogyra (Spirogyre)
Le genre Spirogyra est l'un des plus grands et des plus répandus au monde : on le trouve même en Antarctique. Un fossé, une mare, un étang ou un lac rare est dépourvu de mucus au toucher, flottant à la surface de la boue. Le thalle de Spirogyra est filamenteux, non ramifié, toutes les cellules du filament sont équivalentes et du même type. La cellule a un ou plusieurs chromatophores pariétaux, en spirale, en forme de ruban, avec un grand nombre de pyrénoïdes situés le long de l'axe longitudinal. Les bords du chromatophore sont inégaux. La cellule a une ou plusieurs vacuoles avec la sève cellulaire. Dans le premier cas, le cytoplasme occupe une position maigre. En présence de plusieurs vacuoles, en plus de la couche pariétale du cytoplasme, il existe des cordons cytoplasmiques et un sac cytoplasmique central, dans lequel se trouve un gros noyau clairement visible sans couleur. La couche interne de la membrane cellulaire est la cellulose, la couche externe est la pectine, qui fournit du mucus et la formation d'une couverture gélatineuse, qui donne aux fils un aspect soyeux. Chez les spirogyres, vivant dans des réservoirs à fort courant, différents types de rhizoïdes sont produits qui maintiennent les algues en place. Spirogyra, comme d'autres conjugués, n'a pas de stade flagellaire dans le cycle de développement et ne forme pas de spores. Il se reproduit soit par fragmentation des fils végétativement soit sexuellement. Au cours de la conjugaison, de nombreux zygotes se forment qui, après une période de dormance, germent avec un fil. Sur les quatre noyaux haploïdes formés au cours du processus de fission par réduction, trois petits meurent et il reste un gros noyau viable. Tous les nutriments accumulés dans le zygote vont à la formation d'un plant. Les zygotes sont bien adaptés pour supporter des conditions défavorables. Ils sont recouverts d'une épaisse coquille sculptée à trois couches. La structure de la coquille du zygote est une caractéristique taxonomique importante.
Objets : r. spirogyre.
Progrès
Considérez à m. et esquisser l'apparence du thalle multicellulaire de Spirogyra. Étiquetez la cellule du thalle.
1. Considérez en b. emmené. et dessinez une cage à spirogyra. Nommez la membrane cellulaire, le chromatophore, les pyrénoïdes,
noyau, brins de cytoplasme.
2. Observez et dessinez les différentes étapes du processus de conjugaison.
Désignez les étapes: formation de processus conjugatifs, contraction et débordement du protoplaste, formation du zygote.
Commander Desmidia (Desmidiales)
Ordre Desmidia - organismes unicellulaires ou colonies filamenteuses. La cellule se compose de deux moitiés égales - des demi-cellules. Lors de la propagation végétative, chaque demi-cellule achève la construction de la seconde moitié.
Baguette Closterium (clostérium)
Klosterium est une algue benthique d'eau douce qui nécessite un bon éclairage, elle vit dans de petits réservoirs, des étangs, des marigots calmes de rivières et dans l'encrassement d'objets sous-marins. Avec le développement de masse, des accumulations de mucus se forment. Klosterium aime l'eau propre, mais tolère la pollution organique et se retrouve parfois dans les eaux usées. C'est une forme unicellulaire, son corps en forme de faucille se compose de deux moitiés symétriques - des demi-cellules. Le noyau est situé au centre, dans le sac cytoplasmique. Closterium n'a pas de constriction externe caractéristique des autres algues desmidiées, mais la structure interne correspond aux caractéristiques des représentants de cet ordre. Closterium a deux chromatophores axiaux identiques, une structure particulière. Plusieurs plaques s'étendent radialement à partir de la tige centrale de sorte que le chromatophore ressemble à une étoile à faisceaux multiples en coupe transversale. De grands pyrénoïdes sont situés le long de l'arbre ou dispersés au hasard à la base. La base du chromatophore, tournée vers le centre de la cellule, est large ; aux extrémités de la cellule, le chromatophore se rétrécit coniquement. Aux pôles de la cellule, il y a deux petites vacuoles avec de la sève cellulaire, dans lesquelles sont immergés de petits cristaux de gypse, qui sont en mouvement constant (mouvement brownien), et des corps muqueux spéciaux. La coquille à trois couches du closterium est imprégnée de nombreux pores coniques. Des pores particulièrement grands sont situés aux extrémités des cellules. Ces pores sécrètent du mucus, grâce auquel l'algue se déplace lentement. Alors qu'une extrémité du corps est attachée au substrat, l'autre extrémité oscille. L'algue est ensuite attachée au substrat avec l'autre extrémité. Ainsi culbutant, le closterium se dirige vers la source lumineuse.
La reproduction de Closterium est végétative. Les cellules se divisent transversalement. Chaque cellule fille reçoit la moitié de la cellule mère avec un chromatophore. La seconde moitié, c'est-à-dire la demi-cellule, est en train d'être complétée à nouveau. Premièrement, la jeune demi-cellule n'a pas de chloroplaste, et alors seulement le chromatophore de l'ancienne demi-cellule se divise et la moitié passe dans une nouvelle demi-cellule. Ainsi, chaque individu du closterium est constitué de deux moitiés d'âges différents : l'une est plus âgée et l'autre est plus jeune. Le processus sexuel est la conjugaison de deux individus immergés dans un mucus commun. Le zygote est recouvert d'une membrane en couches épaisses et est bien adapté pour supporter des conditions défavorables. Tout l'hiver, les zygotes sont au repos, et pendant longtemps les noyaux restent non fusionnés. A partir du zygote, deux jeunes clostériums se forment, recevant chacun deux noyaux méiotiques haploïdes. L'un, petit, dégénère bientôt. Le second devient le noyau d'un nouvel individu. Le cycle de développement est monohaplobionte.
Objets : r. clostérium.
Progrès
1. Considérez le closterium à m. et regardez-le bouger.
2. Examinez la cellule de closterium en b. emmené. et dessinez-le. Désigner la membrane cellulaire, le noyau, les chromatophores, les pyrénoïdes, les vacuoles terminales avec des cristaux de gypse.
Questions et tâches
1. Lister les principaux types d'organisation morphologique des algues vertes. Décrivez chaque type. Donne des exemples.
2. Quels signes sont typiques pour les représentants du département des algues vertes ?
3. En quelles classes le département des algues vertes est-il divisé ? Quelle est la différence entre les représentants de chaque classe ?
4. Que sont les colonies et les coenobia chez les algues vertes ? Donnez des exemples d'algues vertes coloniales et cénobiennes. Quelle est la différence entre les organismes coloniaux et multicellulaires ?
5. Comment se déroule la reproduction végétative chez les algues vertes ? Donne des exemples.
6. Comment se produit la reproduction asexuée chez les algues vertes ? Donne des exemples.
7. Quels processus sexuels sont caractéristiques des algues vertes ? Donne des exemples.
8. Dessinez schématiquement le cycle de vie de chlamydomonda, ulotrix, ulva, spirogyra. Inscrivez sur le diagramme quelle génération est un sporophyte et laquelle est un gamétophyte, un ensemble de chromosomes (haploïdes ou diploïdes) pour chaque génération et des cellules utilisées pour la reproduction, le type de division de réduction et le processus sexuel. Indiquez le type de cycle de vie typique de ces algues.
9. Remplissez le tableau :
Caractéristiques comparées des classes et des ordres
10. Décrire le rôle des algues vertes dans la vie des plans d'eau. Donnez des exemples d'algues vertes qui mènent une existence terrestre.
Département des Diatomées (Bacillariophytes, Diatomées)
Un vaste département d'organismes unicellulaires et coloniaux, réunissant plus de 10 000 espèces. Les cellules de diatomées sont recouvertes d'une coquille de silice, composée de deux moitiés qui s'emboîtent l'une sur l'autre, comme un couvercle sur une boîte. La plus grande moitié est l'épithèque et la plus petite moitié est l'hypothèque. Chaque moitié se compose d'une ceinture (en bas) et d'un anneau de ceinture (ceinture) soudés dessus. De plus, la ceinture de l'épithèque se superpose à la ceinture de l'hypothèque. Sous la coquille se trouve une coquille de pectine. Les diatomées appartiennent au groupe des algues de couleur brune, qui se caractérisent par la présence de chlorophylles. un et Avec, masqué par le pigment jaune fucoxanthine. Le produit de réserve de nature glucidique est la chrysolaminarine. Les cellules monadiques sont des spermatozoïdes avec un flagelle penné. Les diatomées se reproduisent végétativement, par division cellulaire longitudinale avec l'achèvement d'une valve - hypothèque. Processus sexuels - conjugaison et oogamie. Après le processus sexuel, un zygote est formé qui a la capacité de se développer (auxospore). Les diatomées vivent à l'état diploïde et seuls leurs gamètes sont haploïdes.
Centré sur la classe (Centrophycées)
La classe Centric combine des algues avec une coquille à symétrie radiale et l'absence d'appareil suture-nodulaire. Toutes les algues centrées sont immobiles. Le processus sexuel est l'oogamie.
Rod Melozira (Mélosire)
Melozira est une algue coloniale filamenteuse, constituée du même type de cellules cylindriques, reliées entre elles par le contact de petites épines situées sur les surfaces arrondies des valves. La coquille du melozira a de larges ceintures, par conséquent, le plus souvent, les algues sont vues de sa surface latérale. Les cellules ont plusieurs chromatophores lobés situés le long de la paroi, le centre de la cellule est occupé par une grande vacuole avec du jus cellulaire.
Melozira n'a pas de structure suture-nodale et est donc immobile.
Se reproduit par division et sexuée. Le processus sexuel est l'oogamie. Dans certaines cellules, après division par réduction, un ovule se forme, dans d'autres - quatre spermatozoïdes avec un flagelle. Le zygote est recouvert d'une fine membrane de pectine bien extensible. Les zygotes à croissance intensive sont appelés auxospores. Puisque les cellules écrasées après des divisions répétées participent au processus sexuel, l'auxospore restaure leur volume d'origine. Ayant atteint une certaine taille, l'auxospore développe sa propre coquille. Le cycle de développement de Melosyra est monodiplobiont.
Objets : r. mélozira.
Progrès
1. Examiner le site de la colonie Melosyra à b. emmené. et esquissez les algues du côté de la ceinture et du côté de la ceinture. Étiquetez la cellule de la colonie, l'épithèque, l'hypothèque.
2. Localisez et dessinez l'auxospore.
Pennate de classe (Pennatophycées)
La classe Pennate comprend les algues à symétrie de coquille bilatérale, ayant un appareil suture-nodulaire et ayant la capacité de se déplacer. Le processus sexuel est la conjugaison.
Genre Pinnularia (Pinnulaire)
Le genre Pinnularia comprend plus de 150 espèces. Il vit dans des plans d'eau douce et pauvre en chaux. Mène un mode de vie benthique au fond ou dans l'encrassement des objets sous-marins. Pinnularia, comme d'autres diatomées, est d'une grande importance en tant que base alimentaire pour les petits animaux et constitue le premier maillon des chaînes alimentaires dans les écosystèmes aquatiques. Il s'agit d'une algue unicellulaire avec une structure suture-nodulaire et, par conséquent, est mobile. Parmi les autres diatomées unicellulaires, la pinnulaire est grande et donc pratique à étudier. À partir de la ceinture, la coquille a un contour rectangulaire et les valves sont linéaires à elliptiques. Les extrémités des valves sont pour la plupart arrondies, mais peuvent être atténuées et capitées. Des nodules et deux ouvertures en forme de fente en forme de S (suture) s'étendant des nodules périphériques au nodule central sont clairement visibles au centre et aux extrémités de la valve. Le long des bords de la ceinture est clairement visible, en particulier sur les coquilles vides, un motif clair de cloisons parallèles - septembre, n'atteignant pas la couture. Les cellules de Pinnularia sont mononucléaires avec deux chromatophores lamellaires, avec des bords incurvés. Le large côté plat du chromatophore fait face au côté de la ceinture et, avec ses bords, il va du côté de la valve. Les cellules actives vivantes de Pinnularia sont colorées en brun jaunâtre, car la fucoxanthine masque les pigments verts, mais dans les cellules mourantes, la fucoxanthine est lessivée et le chromatophore devient vert. Les cellules ont deux vacuoles séparées par un pont cytoplasmique central. Il contient le noyau. Pinnularia stocke la volutine, visible au microscope optique sous forme de corps sphériques faiblement brillants et de gouttes d'huile. Sous la coquille, la cellule est habillée d'une membrane de pectine mucilagineuse. Pinnularia, qui a un appareil suture-nodulaire, se déplace activement, rampant le long du substrat.
Pinnularia se reproduit par division, parallèlement aux valves. Chaque valvule enfant reçoit une valvule mère, tandis que la seconde, valvule complétée, est toujours une hypothèque. En raison de cette caractéristique, à chaque division, une cellule fille est toujours un peu plus petite que la cellule mère, et des individus de tailles différentes peuvent être trouvés dans la population. Le processus sexuel chez Pinnularia n'a pas été trouvé, les auxospores ne se forment pas. On peut supposer que les grandes cellules se divisent plus souvent que les petites et que les plus petites ne se divisent pas du tout. Le cycle de vie est monodiplobiont.
Objet : r. pinnulaire.
Progrès
1. Considérez la pinnularia en b. emmené. Dessinez la cage du côté de la ceinture. Désigner la membrane cellulaire, la suture, les nodules, les septa, le chromatophore.
2. Dessinez la cage à partir du côté de la ceinture. Étiquetez l'épithèque et l'hypothèque.
Questions et tâches
1. Quels niveaux et types d'organisation morphologique sont typiques des représentants des diatomées ?
2. Quelles sont les caractéristiques structurelles des cellules de diatomées ?
3. Quelle est la structure de la coquille des diatomées ?
4. Quels sont les principes de classification des diatomées ?
5. Quelles sont les méthodes de reproduction typiques des diatomées ?
6. Comment se déroulent les processus sexuels chez les diatomées ? Qu'est-ce qu'une auxospore ?
7. Où vivent les diatomées ? Quelles sont les caractéristiques d'adaptation au mode de vie planctonique et benthique des diatomées ? Donnez des exemples de diatomées planctoniques et benthiques.
8. Esquissez schématiquement le cycle de vie des pinnularia et melosira. Inscrivez sur le diagramme quelle génération est un sporophyte et laquelle est un gamétophyte, un ensemble de chromosomes (haploïdes ou diploïdes) pour chaque génération et des cellules utilisées pour la reproduction, le type de division de réduction et le processus sexuel. Indiquez le type de cycle de vie typique de ces algues.
Département Algues brunes (Phéophyte)
Les principaux pigments des algues brunes sont les chlorophylles. un et Avec, des caroténoïdes et des xanthophylles, dont la fucoxanthine, qui masque les pigments verts et donne aux algues une couleur brune caractéristique. Produits de rechange - laminarine (substance de nature glucidique); le mannitol est un alcool de sucre et une petite quantité de graisse. Les phéoplastes sont lamellaires ou plus souvent nombreux discoïdes (chromatophore granuleux) situés dans l'espace périnucléaire sous la membrane nucléaire externe, qui recouvre chaque phéoplaste, formant un réticulum endoplasmique phéoplastique. Les pyrénoïdes dépassent de la surface sous la forme d'un rein. Les cellules ont un noyau, de grandes vacuoles contenant du jus cellulaire et de petites vacuoles contenant des tanins et appelées physodes.
La coquille se compose de deux couches: l'extérieure mucilagineuse, contenant de l'acide alginique, et l'intérieure, formée d'un type spécial de cellulose - l'alguose. Les cellules monadiques avec deux flagelles hétérocontes et hétéromorphes sont situées sur le côté latéral de la cellule. La partie antérieure est longue pennée, couverte de mastigonèmes, la partie postérieure est courte et lisse. L'éventail des structures morphologiques est large : des hétérotriches filamenteux aux formes tissulaires lamellaires différenciées. Chez toutes les algues brunes, à l'exception des représentants de l'ordre des Fucus, dépourvus de reproduction asexuée et diplobiontes, on observe un changement de génération : chez certaines, il est isomorphe, chez d'autres, il est hétéromorphe. Ces différents types de cycle de vie constituent la base de la division moderne des algues brunes en trois classes.
À quelques exceptions près, les algues brunes sont des algues marines, particulièrement abondantes dans les eaux froides des hémisphères nord et sud.
Classe isogénérée (Isogénératophycées)
La classe Isogenate comprend les algues à alternance isomorphe de générations ou à hétéromorphe, mais avec la dominance du gamétophyte.
Ordre Ectocarpe (Ectocarpales) comprend les formes hétérotriches.
Genre Ectocarpus (Ectocarpe)
Ectocarpus est une algue marine benthique répandue, présente sous toutes les latitudes de la bande côtière des mers et des océans. Les algues se déposent sur les rochers et autres objets sous-marins, y compris les plantes. Tolérant à différentes salinités de l'eau, les ectocarpes participent à l'encrassement des navires. La plage de température des habitats d'ectocarpus est également large. Il pousse à la fois dans les mers froides et chaudes et est actif en été comme en hiver. L'ectocarpe est une algue macroscopique (jusqu'à 60 cm), hétérotriche filamenteuse, ayant la forme de buissons ramifiés.
À la base se trouvent des rhizoïdes rampants horizontaux qui fixent les algues au substrat. Les branches verticales à la base du thalle, recouvertes d'une écorce de filaments rhizoïdaux, s'amincissent vers l'apex et se terminent par de longues cellules incolores. L'augmentation est due à la division des cellules situées dans différentes parties du thalle. Dans les cellules de l'ectocarpe, il existe de petites vacuoles avec du jus cellulaire, un noyau dans la couche pariétale du cytoplasme et plusieurs chromatophores en forme de ruban avec des pyrénoïdes. Dans les cellules sénescentes, le chromatophore devient en forme de disque.
La reproduction de l'ectocarpe est asexuée et sexuée. Les méiozoospores, en forme de rein avec deux flagelles inégaux attachés à la surface latérale, sont formées dans des sporanges unicellulaires assis sur une tige unicellulaire. Ces sporanges sont formés sur des plantes sporophytes diploïdes. Sur les plantes haploïdes, des organes reproducteurs multi-nichés sont pondus. Chaque nid produit une cellule reproductrice de l'organisation monadique. Le plus souvent, ces cellules se comportent comme des gamètes et fusionnent pour former un zygote. Ils sont morphologiquement du même type, mais leur comportement diffère : physiologiquement, les gamètes femelles sont moins mobiles et se déposent rapidement au fond ; physiologiquement les hommes sont plus actifs. Le zygote se développe en un sporophyte diploïde. Le cycle décrit ci-dessus correspond au type diplo-haplobionte avec alternance isomorphe des générations, mais l'ectocarpe présente des déviations par rapport à ce cycle, par exemple, les gamètes peuvent germer parthénogénétiquement sans fécondation, donnant de nouveaux individus haploïdes. Ainsi, apparemment, l'éventuelle adaptation complète de l'ectocarpe aux conditions d'existence est réalisée et sa large amplitude écologique est expliquée.
Objets : r. ectocarpus (spécimens d'herbier, micropréparations).
Progrès
1. Considérez les spécimens d'herbier de l'ectocarpe. Dessinez l'apparence du thalle hétérotricheux de l'ectocarpe.
2. Considérez en b. emmené. et puiser dans les sections de préparation du thalle de l'ectocarpe avec des gamétanges à plusieurs chambres et des sporanges à chambre unique. Nommez les cellules du thalle, des gamétanges, des sporanges.
Classe hétérogénérée (Hétérogénératophycées)
La classe hétérogène se caractérise par une alternance hétéromorphe de générations à dominance sporophytique et de petits gamétophytes microscopiques.
Comprend les algues avec un thalle complexe ayant une structure tissulaire. La croissance est réalisée en raison de la division des cellules situées au point de transition de la partie en forme de feuille du thalle vers le pétiole (croissance intercalaire).
Genre Laminaria (Laminaire)
Deux types de varech sont connus et répandus dans les mers du nord : L. sucrier (L. saccharina (L.) Lamour) et L. palmé (L. digitata (Hudz. Lam)), qui forment des plantations entières dans le sublittoral supérieur et sont des algues commerciales. La laminaire est appelée "algue" et est un produit alimentaire précieux utilisé dans l'alimentation animale, ainsi que dans la fabrication de divers produits alimentaires et médicaments pour l'homme.
Laminaria est une grande algue benthique attachée, différenciée de manière complexe, avec une véritable structure tissulaire. Ses cellules méristématiques sont capables de se diviser dans trois directions mutuellement perpendiculaires et de former des structures tridimensionnelles dans lesquelles toutes les cellules sont reliées les unes aux autres par des plasmodesmes. Le thalle du varech se différencie en trois parties clairement distinguables: de puissants rhizoïdes en forme de griffes avec lesquels il se fixe au substrat, une partie à symétrie radiale, appelée pétiole, et une plaque aplatie. Les deux premières structures sont pérennes, tandis que le limbe meurt et repousse à cause du méristème intercalaire situé dans la partie supérieure du pétiole. L'organisation interne du corps du varech est également assez complexe. Le pétiole de la surface a des cellules densément fermées contenant des grains de phéoplastes. Dans les couches plus profondes, les cellules sont incolores et allongées dans le sens longitudinal. Au centre de la cellule sont situés lâchement et forment le noyau. Le pétiole grossit progressivement et des couches ressemblant à des anneaux de croissance sont clairement visibles sur la coupe transversale. Dans la structure anatomique de la plaque, on distingue également le cortex à petites cellules. Au centre se trouve une "veine", dont les cellules ressemblent dans leur structure aux cellules criblées des plantes supérieures.
Les laminaires se reproduisent sexuellement et asexuellement. Les sporanges sont situés à la surface de la plaque, formant des zones ou des champs entiers porteurs de spores, sur lesquels des sporanges en forme de sac et des processus paraphysaires stériles allongés sont situés dans une couche de palissade. Dans les sporanges, après division par réduction, des méiozoospores réniformes biflagellées se forment et germent en organismes haploïdes. Les gamétophytes femelles et mâles diffèrent du sporophyte. Ce sont des plantes microscopiques filamenteuses et sous-développées (excroissances). Ils sont de courte durée, leur fonction principale est la production de gamètes. Le processus sexuel est l'oogamie. Le zygote germe sans période de dormance et se développe en un sporophyte diploïde. Ainsi, presque toute la vie active du varech se produit à l'état diploïde. Le cycle de développement est diplohaplobionte avec alternance hétérotrophe de générations, à dominance sporophytique.
Fin du segment d'introduction.
ALGUES : CLASSIFICATION DES ALGUES
Vers l'article ALGUES
Dans le passé, les algues étaient considérées comme des plantes primitives (sans tissus conducteurs ou vasculaires spécialisés) ; ils ont été isolés dans la subdivision des algues (Algae), qui, avec la subdivision des champignons (Fungi), constituait la division du thalle (couche), ou plantes inférieures (Thallophyta), l'une des quatre divisions du règne végétal ( certains auteurs utilisent le terme zoologique au lieu du terme "département" de type"). De plus, les algues ont été divisées par couleur - en vert, rouge, marron, etc. La couleur est assez forte, mais pas la seule base pour classification générale ces organismes. Les types de formation de leurs colonies, les méthodes de reproduction, les caractéristiques des chloroplastes, la paroi cellulaire, les substances de réserve, etc. sont plus essentielles pour la sélection des différents groupes d'algues. Les anciens systèmes reconnaissaient généralement une dizaine de ces groupes, qui étaient considérés comme des classes. L'un des systèmes modernes fait référence aux « algues » (ce terme a perdu sa valeur de classification) huit types (divisions) du royaume protiste (Protista) ; cependant, cette approche n'est pas reconnue par tous les scientifiques.
Les algues vertes constituent la division (phylum) Chlorophyta du règne protiste. Ils sont généralement de la couleur du vert gazon (bien que la couleur puisse varier du jaune pâle au presque noir) et leurs pigments photosynthétiques sont les mêmes que ceux des plantes ordinaires. La plupart sont des formes microscopiques d'eau douce. De nombreuses espèces poussent sur le sol, formant des raids feutrés sur sa surface humide. Ils sont unicellulaires et multicellulaires, forment des filaments, des colonies sphériques, des structures en forme de feuille, etc. Les cellules sont mobiles (avec deux flagelles) ou immobiles. reproduction sexuée - différents niveaux difficulté selon le type. Plusieurs milliers d'espèces ont été décrites. Les cellules contiennent un noyau et plusieurs chloroplastes distincts. L'un des genres bien connus est Pleurococcus, une algue unicellulaire qui forme les taches vertes souvent observées sur l'écorce des arbres. Le genre Spirogyra est répandu - des algues filamenteuses qui forment de longues fibres de boue dans les ruisseaux et les rivières froides. Au printemps, ils flottent en touffes collantes vert jaunâtre à la surface des étangs. Cladophora pousse sous la forme de "buissons" mous et fortement ramifiés qui s'attachent aux pierres le long des berges des rivières. Basiocladia forme un revêtement vert sur le dos des tortues d'eau douce. Le maillage d'eau (Hydrodictyon) composé de nombreuses cellules, vivant dans des eaux stagnantes, ressemble vraiment à un "sac à cordes" par sa structure. Desmidia - algues vertes unicellulaires qui préfèrent l'eau douce des marais; leurs cellules se distinguent par une forme bizarre et une surface magnifiquement ornée. Chez certaines espèces, les cellules sont connectées en colonies filamenteuses. Dans l'algue coloniale flottante Scenedesmus, les cellules en forme de faucille ou oblongues sont disposées en courtes chaînes. Ce genre est courant dans les aquariums, où sa reproduction en masse entraîne l'apparition d'un "brouillard" vert dans l'eau. La plus grande algue verte est la laitue de mer (Ulva), un macrophyte en forme de feuille.
Les algues rouges (cramoisies) constituent le département (type) Rhodophyta du règne protiste. La plupart d'entre eux sont des macrophytes feuillus marins, buissonnants ou croustillants vivant sous la ligne de marée basse. Leur couleur est majoritairement rouge en raison de la présence du pigment phycoérythrine, mais peut être violette ou bleutée. Certains violets se trouvent dans l'eau douce, principalement dans les ruisseaux et les rivières rapides et claires. Batrachospermum est une algue gélatineuse très ramifiée composée de cellules brunâtres ou rougeâtres ressemblant à des perles. Lemanea est une forme en forme de brosse qui pousse souvent dans les ruisseaux et les cascades à débit rapide où ses thalles se fixent aux rochers. Audouinella est une algue filamenteuse présente dans les petites rivières. La mousse d'Irlande (Chondrus cripus) est un macrophyte marin commun. Les violets ne forment pas de cellules mobiles. Leur processus sexuel est très complexe et un cycle de vie comprend plusieurs phases.
Les algues brunes constituent la division Phaeophyta (type) du royaume des protistes. Presque tous sont des habitants de la mer. Seules quelques espèces sont microscopiques, et parmi les macrophytes se trouvent les plus grandes algues du monde. Ce dernier groupe comprend le varech, les macrocystis, les fucus, les sargasses et les lessonia ("palmiers marins"), les plus abondants le long des côtes des mers froides. Toutes les algues brunes sont multicellulaires. Leur couleur varie du jaune verdâtre au brun foncé et est due au pigment fucoxanthine. La reproduction sexuée est associée à la formation de gamètes mobiles à deux flagelles latéraux. Les instances qui forment des gamètes sont souvent complètement différentes des organismes de la même espèce qui se reproduisent uniquement par des spores.
Les diatomées (diatomées) sont regroupées dans la classe Bacillariophyceae, qui, dans la classification utilisée ici, est incluse, avec les algues dorées et jaune-vert, dans le département (type) Chrysophyta du royaume des protistes. Les diatomées sont un très grand groupe d'espèces marines et d'eau douce unicellulaires. Leur couleur va du jaune au brun en raison de la présence du pigment fucoxanthine. Le protoplaste des diatomées est protégé par une coquille de silice (verre) en forme de boîte - une coquille composée de deux valves. La surface dure des valves est souvent recouverte d'un motif complexe de stries, de tubercules, de creux et de crêtes caractéristiques de l'espèce. Ces coquilles sont l'un des plus beaux objets microscopiques, et la clarté de la distinction de leurs motifs est parfois utilisée pour tester le pouvoir de résolution d'un microscope. Habituellement, les valves sont percées de pores ou présentent un espace appelé couture. La cellule contient le noyau. Outre la division cellulaire en deux, la reproduction sexuée est également connue. De nombreuses diatomées sont des formes nageant librement, mais certaines sont attachées à des objets sous-marins avec des tiges visqueuses. Parfois, les cellules sont combinées en fils, chaînes ou colonies. Il existe deux types de diatomées: les cirrus avec des cellules allongées à symétrie bilatérale (elles sont plus abondantes dans les eaux douces) et centriques, dont les cellules, vues de la valve, semblent arrondies ou polygonales (elles sont plus abondantes dans les mers).
Comme déjà mentionné, les coquilles de ces algues persistent après la mort cellulaire et se déposent au fond des plans d'eau. Au fil du temps, leurs puissantes accumulations se compactent en une couche poreuse rock- diatomite.
Flagelles. Ces organismes, en raison de leur capacité à la nutrition "animale" et d'un certain nombre d'autres caractéristiques importantes, sont maintenant souvent appelés le sous-royaume des protozoaires (Protozoaires) du royaume des protistes, mais ils peuvent également être considérés comme une division (type) d'Euglenophyta du même royaume, qui n'est pas inclus dans les Protozoaires. Tous les flagelles sont unicellulaires et mobiles. Les cellules sont vertes, rouges ou incolores. Certaines espèces sont capables de photosynthèse, tandis que d'autres (saprophytes) absorbent la matière organique dissoute ou même avalent ses particules solides. La reproduction sexuée n'est connue que chez certaines espèces. Un habitant commun des étangs est Euglena, une algue verte à œil rouge. Elle nage à l'aide d'un seul flagelle, est capable à la fois de photosynthèse et de nutrition de matière organique prête à l'emploi. Euglena sanguinea peut rendre l'eau de l'étang rouge à la fin de l'été.
Dinoflagellés. Ces organismes flagellaires unicellulaires sont également souvent appelés protozoaires, mais ils peuvent également être distingués en tant que département indépendant (type) Pyrrophyta du royaume protiste. Ils sont principalement jaune-brun, mais ils peuvent aussi être incolores. Leurs cellules sont généralement mobiles ; la paroi cellulaire est absente chez certaines espèces, et parfois elle a une forme très bizarre. La reproduction sexuée n'est connue que chez quelques espèces. Le genre marin Gonyaulax est l'une des causes des « marées rouges » : près des côtes, il est si abondant que l'eau prend une couleur inhabituelle. Cette algue libère des substances toxiques, entraînant parfois la mort de poissons et crustacés. Certains dinoflagellés provoquent la phosphorescence de l'eau dans les mers tropicales.
Les algues dorées sont incluses, avec d'autres, dans la division Chrysophyta (type) du royaume protiste. Leur couleur est jaune-brun et les cellules sont mobiles (flagellées) ou immobiles. La reproduction est asexuée avec formation de kystes imprégnés de silice.
Les algues jaune-vert sont maintenant généralement combinées avec des algues dorées dans la division (type) Chrysophyta, mais elles peuvent également être considérées comme une division indépendante (type) Xanthophyta du royaume protiste. Dans la forme, ils ressemblent aux algues vertes, mais diffèrent par la prédominance de pigments jaunes spécifiques. Leurs parois cellulaires sont parfois constituées de deux moitiés entrant l'une dans l'autre, et chez les espèces filamenteuses, ces valves sont en forme de H en coupe longitudinale. La reproduction sexuée n'est connue que sous quelques formes.
Les Charovye (raies) sont des algues multicellulaires qui composent la division Charophyta (type) du royaume des protistes. Leur couleur varie du vert grisâtre au gris. Les parois cellulaires sont souvent incrustées de carbonate de calcium, de sorte que les restes morts d'ombles sont impliqués dans la formation de dépôts de marne. Ces algues ont un axe principal cylindrique en forme de tige, à partir duquel des processus latéraux s'étendent en verticilles, semblables aux feuilles des plantes. Les characées poussent verticalement dans les eaux peu profondes, atteignant une hauteur de 2,5 à 10 cm Reproduction sexuée. Il est peu probable que les characées soient proches de l'un des groupes énumérés ci-dessus, bien que certains botanistes pensent qu'elles descendent d'algues vertes. Voir aussi SYSTÉMATIQUE VÉGÉTALE.
Charbonnier. Dictionnaire de Collier. 2012
Voir aussi les interprétations, les synonymes, les significations du mot et ce qu'est ALGUE : CLASSIFICATION DES ALGUES en russe dans les dictionnaires, encyclopédies et ouvrages de référence :
- ALGUE dans l'Encyclopédie de biologie :
, un vaste groupe d'organismes photosynthétiques, parfois attribués à un règne végétal distinct. Comprend 12 divisions (algues bleu-vert, algues brunes, algues vertes, ... - ALGUE
(Algues) - organismes végétaux inférieurs, classés comme un sous-royaume des spores ou plantes cryptogames (Sporophyta s. Kryptogamae). Avec des champignons et du lichen ... - ALGUE
(algues)? organismes végétaux inférieurs classés comme un sous-royaume des plantes porteuses de spores ou cryptogames (Sporophyta s. Kryptogamae). Avec des champignons et du lichen ... - ALGUE dans le dictionnaire de Collier :
(Algues), un groupe vaste et hétérogène d'organismes primitifs ressemblant à des plantes. À quelques exceptions près, ils contiennent le pigment vert chlorophylle, essentiel pour... - CLASSIFICATION dans le plus récent dictionnaire philosophique :
(lat. classis - catégorie, classe et facio - je fais, agencer) - une division en plusieurs étapes du volume logique d'un concept (logique) ou de tout ... - CLASSIFICATION
MARCHANDISES - cession par les autorités douanières Fédération Russe marchandises spécifiques aux articles spécifiés dans la Nomenclature des marchandises pour l'activité économique extérieure (TN VED). … - CLASSIFICATION dans le Dictionnaire des termes économiques :
IMMOBILISATIONS - un groupement d'immobilisations établi par le Comité national des statistiques de l'URSS et comprenant 12 types : bâtiments ; ouvrages d'art ; dispositifs de transmission ; voitures et équipement; le transport … - CLASSIFICATION dans le Dictionnaire des termes économiques :
BUDGET - voir CLASSIFICATION BUDGETAIRE ... - CLASSIFICATION dans le Dictionnaire des termes économiques :
- distribution, espacement des objets, concepts, noms en classes, groupes, catégories, dans lesquels les objets avec c.-l. tombent dans un groupe. commun ... - CLASSIFICATION dans l'Encyclopédie de biologie :
en biologie, répartition de la diversité des organismes vivants dans un certain ordre selon un système. La classification est basée sur un ensemble de fonctionnalités qui permettent ... - ALGUE dans l'Encyclopédie biblique de Nicéphore :
ou SEA GRASS (Jon 2:6) mer, herbe d'eau, comme lu dans la traduction russe dans la citation indiquée. A été embrassé par l'herbe marine ... - ALGUE en termes médicaux :
(algues) un groupe de plantes inférieures autotrophes porteuses de chlorophylle, généralement aquatiques, non divisées en racines, tiges et feuilles, capables d'absorber le dioxyde de carbone dans ... - CLASSIFICATION
(de lat. classis - classe de catégorie et ... fication), en logique - un système de concepts subordonnés (classes d'objets) de tout domaine de connaissance ou ... - ALGUE dans le Grand Dictionnaire Encyclopédique :
un groupe de plantes aquatiques inférieures qui contiennent généralement de la chlorophylle et produisent de la matière organique par photosynthèse. Le corps de l'algue est un thalle qui n'a pas ... - ALGUE dans la Grande Encyclopédie soviétique, TSB :
(Algues), un groupe de plantes inférieures, autotrophes, généralement aquatiques; contiennent de la chlorophylle et d'autres pigments et produisent de la matière organique par photosynthèse. … - CLASSIFICATION dans le dictionnaire encyclopédique de Brockhaus et Euphron :
un dispositif logique très important qui est utilisé dans l'étude du sujet et qui est basé sur la division logique des concepts. En effet, le classement n'est pas... - CLASSIFICATION dans le dictionnaire encyclopédique moderne :
- CLASSIFICATION
(du latin classis - catégorie, classe et ... fication) (en logique), un système de concepts subordonnés (classes d'objets) de tout domaine de connaissance ou d'activité ... - CLASSIFICATION dans le dictionnaire encyclopédique :
et bien. 1. pl. non. La répartition de certains objets en classes en fonction de leurs propriétés.||Cf. RUBRICATION. 2. … - CLASSIFICATION dans le dictionnaire encyclopédique :
, -si. 1. voir classer. 2. Système, selon un essaim de quelque chose. .classifié. K.Sciences. Bibliothèque K. II adj. classement, e, ... - CLASSIFICATION
CLASSIFICATION DES LANGUES, l'étude et le regroupement des langues du monde de diverses manières. caractéristiques : génétique KI (généalogique) - sur la base de la parenté, c'est-à-dire origine commune... - CLASSIFICATION dans le grand dictionnaire encyclopédique russe :
CLASSIFICATION DES SCIENCES, divulgation de l'interconnexion des sciences sur la base d'une définition. principes (objectif, subjectif, coordination, subordination, etc.) et l'expression de leur connexion ... - CLASSIFICATION dans le grand dictionnaire encyclopédique russe :
CLASSIFICATION (foyer), séparation des particules de minéraux broyés en produits homogènes (classes) en termes de taille, de densité et d'autres produits. K. est produit en ... - CLASSIFICATION dans le grand dictionnaire encyclopédique russe :
CLASSIFICATION (de lat. classis - catégorie, classe et ... fication) (en logique), un système de concepts subordonnés (classes d'objets) c.-l. domaines… - ALGUE dans le grand dictionnaire encyclopédique russe :
ALGUES, groupe preim. l'eau. organismes, contenant généralement de la chlorophylle et produisant de la matière organique. in-va dans le processus de photosynthèse. Le corps de V. est un thalle qui n'a pas ... - CLASSIFICATION dans l'Encyclopédie de Brockhaus et Efron :
? un dispositif logique très important qui est utilisé dans l'étude du sujet et qui est basé sur la division logique des concepts. En effet, le classement est... - CLASSIFICATION
classement, classement, classement, classement, classement, classement, classement, classement, classement, classement, classement, classement, classement, classement, ... - ALGUE dans le paradigme Full accentué selon Zaliznyak :
grandir, grandir, grandir, grandir, grandir, grandir... - CLASSIFICATION dans le Dictionnaire des termes linguistiques :
(de lat. classis - rang + facere - mettre) voyelles, voir les sons des voyelles ... - CLASSIFICATION dans le Dictionnaire populaire explicatif-encyclopédique de la langue russe :
-et bien. 1) Le système de concepts subordonnés (classes d'objets) dans certains branche de la connaissance, compilée sur la base de la prise en compte des propriétés des objets et régulière ... - CLASSIFICATION dans le Thésaurus du vocabulaire commercial russe :
- CLASSIFICATION dans le nouveau dictionnaire des mots étrangers :
(lat.; voir classer) 1) un système de concepts subordonnés (classes d'objets, phénomènes) dans certains. branche de la connaissance, compilée sur la base de ... - CLASSIFICATION dans le Dictionnaire des expressions étrangères :
[lat. ; voir classer] 1. un système de concepts subordonnés (classes d'objets, phénomènes) chez certains. branche de connaissances, compilée sur la base de la prise en compte de points communs ... - CLASSIFICATION dans le thésaurus russe :
Syn : classification, systématisation, systématisation, taxonomie, regroupement, tri, placement, ... - CLASSIFICATION dans le Dictionnaire des synonymes d'Abramov :
cm. … - CLASSIFICATION dans le dictionnaire des synonymes de la langue russe :
Syn : classification, systématisation, systématisation, taxonomie, regroupement, tri, placement, ... - CLASSIFICATION dans le Nouveau dictionnaire explicatif et dérivationnel de la langue russe Efremova :
et. 1) Identique à : classement. 2) Le système de distribution de certains. objets ou concepts homogènes par classes, catégories, etc. selon …
Envoyer votre bon travail dans la base de connaissances est simple. Utilisez le formulaire ci-dessous
Les étudiants, les étudiants diplômés, les jeunes scientifiques qui utilisent la base de connaissances dans leurs études et leur travail vous en seront très reconnaissants.
Posté sur http://allbest.ru
1. ALGUES ( ALGUES)
1.1 CARACTERISTIQUES GENERALES
Les algues sont un groupe d'organismes principalement aquatiques. Un trait caractéristique de toutes les algues est que leur corps n'est pas divisé en organes végétatifs (racine, tige, feuille), mais est représenté par un thalle ou un thalle. Pour cette raison, ils sont appelés thalle ou organismes thalle. Contrairement aux plantes supérieures, elles manquent généralement de tissus et les organes de reproduction sexuée sont généralement unicellulaires. Le point commun aux algues est leur capacité à se nourrir en mode autotrophe en raison de la présence d'un appareil photosynthétique. Dans le même temps, chez certaines algues, parallèlement à la nutrition autotrophe, il existe également une nutrition hétérotrophe.
Plus de 40 000 espèces d'algues sont connues, qui sont combinées en 11 divisions: diatomées - environ 20 000 espèces, vertes - 13-20 000, rouges - environ 4 000, bleu-vert - environ 2 000, brunes - environ 1 000, dinophytes et cryptophytes - plus plus de 1 000, jaune-vert, doré, characées - plus de 300 dans chaque division, euglénoïdes - environ 840 espèces. Selon le célèbre algologue biélorusse T.M. Mikheeva (1999) a trouvé 1832 espèces d'algues en Biélorussie, et avec des taxons intraspécifiques - 2338 représentants. Les espèces découvertes appartiennent à 363 genres dans 134 familles de 10 divisions. Dans le même temps, 21 espèces d'algues sont répertoriées dans le Livre rouge de la République de Biélorussie.
Structure d'algues. Les algues au sein de la structure de type thalle se distinguent par une diversité morphologique exceptionnelle. Leur corps peut être unicellulaire, colonial, multicellulaire. Leurs tailles au sein de chacune de ces formes diffèrent dans une large gamme - du microscopique (1 micron) au gigantesque (il existe des espèces atteignant plusieurs dizaines de mètres). Compte tenu de la grande diversité morphologique du corps végétatif, les algues peuvent être divisées en plusieurs catégories selon leur structure, formant les principales étapes de l'évolution morphologique.
La structure monadique (flagellaire) est caractéristique des organismes unicellulaires et coloniaux et se caractérise par la présence de cellules d'un, deux ou plusieurs flagelles, qui déterminent le mouvement actif dans l'eau. Cette structure prévaut chez les dinophytes et les cryptophytes, les algues dorées et euglènes. Chez les algues plus hautement organisées, les cellules qui servent à la reproduction asexuée (zoospores) ou sexuée (gamètes) ont une structure monadique.
La structure amiboïde (rhizopodiale) est caractérisée par l'absence d'une forme cellulaire permanente, d'une membrane dense et de flagelles. Ces algues, comme les amibes, se déplacent à l'aide de pseudopodes, qui sont conservés dans les dinophytes, les algues dorées et jaune-vert.
La structure palmelloïde (hémimonasale ou capsale) est une combinaison de nombreuses cellules immobiles immergées dans un mucus commun, mais sans connexions plasmatiques. La structure palmelloide est largement représentée dans les algues vertes, jaune-vert et dorées ; dans d'autres départements, elle est moins fréquente ou totalement absente.
La structure coccoïde est caractérisée par des cellules immobiles diverses formes et tailles, avec une paroi cellulaire dense, simple ou connectée dans une colonie (cénobie). Une telle structure se retrouve dans presque tous les départements (à l'exception de l'euglène) des algues, et dans les diatomées c'est la seule; chez d'autres représentants, on l'observe dans les cycles de développement (aplanospores, akinètes, tétraspores, etc.).
La structure filamenteuse (trichale) dans le monde des algues est la forme la plus simple thalle multicellulaire et est une connexion de cellules immobiles en fils, entre lesquels une interaction physiologique est réalisée à l'aide de plasmodesmes. Les fils peuvent être simples et ramifiés, libres, attachés et réunis le plus souvent en colonies muqueuses. La structure filamenteuse se présente parmi les algues vertes, dorées, jaune-vert, rouges.
La structure multifilamenteuse (hétérotriche) est une variante plus complexe de la structure filamenteuse, qui se caractérise par deux systèmes de filaments : ceux qui fluent le long du substrat et ceux qui s'étendent verticalement à partir de ceux-ci.
La structure hétérotriche est caractéristique de nombreuses algues bleues-vertes, vertes, carbonisées, dorées, jaune-vertes, rouges et brunes et peut être une forme permanente ou temporaire.
La structure pseudoparenchymateuse (faux tissu) se caractérise par la formation de grands thalles volumineux résultant de la fusion des fils d'un thalle multifilamenteux, parfois accompagnée d'une différenciation des "tissus". Étant donné que ces derniers, en termes de méthode de formation, diffèrent des vrais, ils sont appelés faux tissus. Trouvé dans certaines algues rouges.
La structure siphonale (siphon) est un thalle, souvent de grande taille et de différenciation morphologique complexe, sans cloisons cellulaires et généralement avec de nombreux noyaux. Le type d'organisation siphonale est présent dans certaines algues vertes et jaune-vert.
La structure siphonocladale se retrouve chez certaines algues vertes filamenteuses, qui se caractérisent par une division ségrégative de cellules multinucléées : le protoplaste se décompose en parties arrondies entourées d'une membrane, donnant naissance à de nouveaux segments du thalle.
Structure cellulaire. L'organisation de la cellule de la plupart des algues (sauf celles bleu-vert) diffère peu de l'organisation des cellules typiques des plantes supérieures, mais elle a aussi ses propres caractéristiques. La cellule de la plupart des algues est habillée d'une membrane cellulaire permanente, a un système à deux phases, se compose d'une matrice amorphe, d'hémicellulose ou de pectine, dans laquelle sont immergés des éléments squelettiques fibreux - des microfibrilles. Dans de nombreuses algues, des composants supplémentaires sont déposés: carbonate de calcium (characeae, acetobularia, padina), acide alginique (marron), fer (rouge). Dans la vie d'une cellule végétale, un rôle important est joué par la présence dans l'enveloppe de fractions d'abord de pectine puis de cellulose, qui assurent des fonctions de support et de protection, ainsi que la capacité de s'imprégner et de se développer. La membrane cellulaire peut être entière ou constituée de deux parties ou plus, pénétrée par des pores, et peut porter diverses excroissances. Sous la coquille se trouve le protoplaste, y compris le cytoplasme et le noyau.
Les algues sont le seul groupe où il existe les trois types d'organisation cellulaire : les procaryotes (algues bleu-vert, où il n'y a pas de noyau, leur rôle est joué par le nucléoïde) ; mésocaryote (dinophytes, il y a un noyau, mais primitif) et eucaryote (les algues des autres divisions sont de véritables organismes nucléaires).
Le cytoplasme de la plupart des algues est situé dans une fine couche de paroi, entourant une grande vacuole centrale avec du jus cellulaire. La vacuole est absente dans les cellules des algues bleu-vert et des monades (des vacuoles pulsantes sont notées dans les monades d'eau douce). Dans le cytoplasme des algues eucaryotes, les éléments du réticulum endoplasmique, des ribosomes, des mitochondries, de l'appareil de Golgi, des chromatophores et des noyaux cellulaires se distinguent clairement ; il y a aussi des lysosomes, des peroxysomes, des sphérosomes.
Dans les cellules d'algues (à l'exception de celles bleu-vert), les chromatophores (chloroplastes) sont particulièrement visibles à partir des organites qui, contrairement aux chloroplastes des plantes supérieures, sont de forme, de couleur, de nombre, de structure et d'emplacement dans la cellule. Ils peuvent être en forme de coupe (chlamydomonas), en spirale (spirogyra), lamellaire (diatomées pennées), cylindrique (edogonium). Chez de nombreuses algues, les chromatophores sont nombreux et ressemblent à des grains ou des disques situés dans le cytoplasme pariétal (vert avec une organisation en siphon, marron, rouge). Les chromatophores sont gainés, composés de stroma, de structures lamellaires qui ressemblent à des sacs aplatis et sont appelés thylakoïdes. Ils contiennent des pigments. De plus, la matrice chromatophore contient des ribosomes, de l'ADN, de l'ARN, des granules lipidiques et des inclusions spéciales de pyrénoïdes. Le pyrénoïde est une formation spécifique inhérente à toutes les algues (à l'exception des bleu-vert) et à un petit groupe de mousses.
Reproduction d'algues. La reproduction asexuée chez les algues unicellulaires est réalisée par division cellulaire, chez les algues coloniales et filamenteuses - à la suite de la décomposition des colonies ou des filaments en fragments séparés; dans quelques algues, des organes reproducteurs spéciaux se forment, par exemple des nodules dans les characées, des akinètes (cellules spéciales contenant une grande quantité de substances de réserve et de pigments) dans les verts, etc. Une telle reproduction est souvent appelée végétative.
La reproduction asexuée se produit également au moyen de spores immobiles (aplanospores) ou de zoospores (spores à flagelles), formées par division de protoplastes de cellules ordinaires ou spéciales appelées sporanges. Chez un certain nombre de représentants d'algues vertes, les aplanospores acquièrent parfois toutes les caractéristiques distinctives de cette cellule déjà dans la cellule mère. Dans de tels cas, ils parlent d'autospores. La reproduction au moyen de spores est appelée reproduction asexuée appropriée.
La reproduction sexuée est caractérisée par la présence d'un processus sexuel dont l'une des étapes les plus importantes est la fécondation, c'est-à-dire fusion de cellules sexuelles haploïdes - gamètes. À la suite de la fécondation, un zygote est formé avec une nouvelle combinaison de traits héréditaires, qui devient l'ancêtre d'un nouvel organisme.
Chez les algues, on distingue les formes suivantes du processus sexuel: chologamie - la fusion de deux individus unicellulaires; isogamie - la fusion de gamètes mobiles de structure et de taille identiques; hétérogamie - la fusion de gamètes mobiles de différentes tailles (le plus grand est considéré comme une femme); oogamie - la fusion d'un gros œuf immobile avec un petit gamète mâle mobile - un spermatozoïde ou un immobile, dépourvu de spermatozoïdes (chez les algues rouges); conjugaison - la fusion de protoplastes de cellules non spécialisées. Les gamètes se forment dans des cellules qui ne diffèrent pas des cellules végétatives ou dans des cellules spéciales appelées gamétanges. Les gamétanges contenant un œuf (rarement plusieurs) sont appelées ovogonies, et celles dans lesquelles se forment des spermatozoïdes ou des spermatozoïdes sont appelées anthéridies. Chez les algues primitives, chaque individu est capable de former à la fois des spores et des gamètes, selon la saison et les conditions extérieures ; dans d'autres, les fonctions de reproduction asexuée et sexuée sont exercées par différents individus - sporophytes (forme de spores) et gamétophytes (forme de gamètes). Les principaux types de cycles de vie des algues.
1. Le type haplophase se caractérise par l'absence d'alternance de générations. Toute la vie végétative des algues se déroule dans un état haploïde, c'est-à-dire qu'elles sont des haplontes. Seul le zygote est diploïde, dont la germination s'accompagne d'une division réductrice du noyau (réduction zygotique). Les plantes qui se développent dans ce cas sont haploïdes. Les exemples sont de nombreuses algues vertes (volvox, la plupart des chlorocoques, conjugués) et char.
2. Le type diplophase se distingue par le fait que toute la vie végétative des algues se déroule à l'état diploïde et que la phase haploïde n'est représentée que par les gamètes. Avant leur formation, une division de réduction du noyau se produit (réduction gamétique). Le zygote sans division nucléaire se transforme en un thalle diploïde. Ces algues sont des diplontes. Ce type de développement est caractéristique de nombreuses algues vertes à structure en siphon, de toutes les diatomées et de quelques représentants des algues brunes (ordre Fucal).
3. Le type diplohaplophase se caractérise par le fait que dans les cellules des thalles diploïdes (sporophytes) de nombreuses algues, la division de réduction du noyau précède la formation de zoo- ou aplanospores (réduction sporique). Les spores se développent en plantes haploïdes (gamétophytes) qui ne se reproduisent que sexuellement. Un œuf fécondé - un zygote - germe en une plante diploïde qui porte des organes de reproduction asexuée. Ainsi, dans ces algues, il existe une alternance de formes évolutives (générations) : un sporophyte asexué diploïde et un gamétophyte sexuel haploïde. Les deux générations peuvent ne pas différer en apparence et occuper la même place dans le cycle de développement (changement isomorphe des générations) ou différer fortement dans les caractéristiques morphologiques (changement hétéromorphe des générations). Un changement isomorphe dans les générations est caractéristique d'un certain nombre d'algues vertes (ulves, entéromorphes, cladophores), brunes et la plupart des algues rouges. Un changement hétéromorphe de générations se produit avec une prédominance à la fois du gamétophyte et du sporophyte (caractéristique principalement des algues brunes, moins souvent vertes et rouges).
L'eau est le principal milieu vivant des algues. De plus, des facteurs tels que la lumière, la température, la salinité de l'eau, la composition chimique du substrat, etc., jouent un rôle extrêmement important dans leur vie.Selon les conditions environnementales, les algues forment divers groupes ou communautés (cénoses), dont chacun se caractérise par une composition spécifique plus ou moins certaine.
1.2 MÉTHODES DE COLLECTE, DE CONSERVATION ET D'ÉTUDE DES ALGUES
Les algues peuvent être récoltées du début du printemps à la fin de l'automne, tandis que les algues terrestres peuvent être récoltées dans des zones sans neige tout au long de l'année. Pour les collecter, vous devez prendre des bocaux à large ouverture et des bouchons bien ajustés, un sac pour eux, un couteau, un grattoir pointu, un filet à plancton, une fiole de formol, des boîtes ou des sacs en plastique pour la collecte d'algues terrestres, du papier à lettres pour les étiquettes, un cahier pour les notes, un crayon.
Les méthodes de collecte et d'étude des algues sont déterminées principalement par les caractéristiques écologiques et morphologiques des représentants de divers départements et groupes écologiques. Considérons les principales méthodes de collecte et d'étude des algues de divers plans d'eau à des fins d'études floristiques-systématiques et en partie hydrobiologiques.
Collection de phytoplancton. Le choix de la méthode d'échantillonnage du phytoplancton dépend du type de réservoir, du degré de développement des algues, des objectifs de recherche, des instruments disponibles, de l'équipement, etc. de la concentration préliminaire de microorganismes vivant dans la colonne d'eau. Une de ces méthodes consiste à filtrer l'eau à travers des filets à plancton (description des filets à plancton et autres dispositifs et dispositifs de collecte d'algues).
Lors de la collecte de plancton à partir des couches superficielles d'un réservoir, le filet à plancton est abaissé dans l'eau de sorte que l'ouverture supérieure du filet se trouve à une distance de 5 à 10 cm au-dessus de la surface de l'eau. Un récipient d'un certain volume puise l'eau de la couche superficielle (jusqu'à 15-20 cm de profondeur) et la verse dans le filet, filtrant ainsi 50 à 100 litres d'eau. Dans les grands réservoirs, les échantillons de plancton sont prélevés à partir d'un bateau : un filet à plancton est tiré sur une corde fine derrière un bateau en mouvement pendant 5 à 10 minutes. Pour les collectes verticales de plancton, des filets de conception spéciale sont utilisés. Dans les petites étendues d'eau, des échantillons de plancton peuvent être prélevés sur le rivage en ramassant soigneusement l'eau dans un récipient devant vous et en la filtrant à travers un filet, ou en jetant un filet sur une fine corde dans l'eau et en le retirant avec précaution. Pour la comptabilité quantitative du phytoplancton, le volume des échantillons est constitué par des dispositifs spéciaux - des bathomètres - de différentes conceptions. Le bathomètre du système Rutner a reçu une large application dans la pratique. Sa partie principale est un cylindre en métal ou en plexiglas, d'une capacité de 1 à 5 litres. L'appareil est équipé de couvercles supérieur et inférieur fermant hermétiquement le cylindre. Sous l'eau, le bathomètre est abaissé avec les couvercles ouverts. Lorsque la profondeur requise est atteinte, à la suite d'une forte secousse de la corde, les couvercles ferment les trous du cylindre qui, une fois fermé, est retiré à la surface. L'eau enfermée dans le cylindre est versée dans le récipient préparé par un tuyau de dérivation latéral équipé d'un robinet. Lors de l'étude du phytoplancton des couches superficielles de l'eau, des échantillons sont prélevés sans l'aide d'un bathomètre en puisant de l'eau dans un récipient d'un certain volume.
Collecte de phytobenthos. Pour étudier la composition spécifique du phytobenthos à la surface d'un réservoir, il suffit d'extraire une certaine quantité de sol de fond et de sédiments sur celui-ci. Dans les eaux peu profondes (jusqu'à 0,5 à 1,0 m de profondeur), ceci est réalisé à l'aide d'un tube à essai abaissé au fond ou d'un siphon - un tuyau en caoutchouc avec des tubes en verre aux extrémités, dans lequel le limon est aspiré. En profondeur, des échantillons de qualité sont prélevés à l'aide d'un seau ou d'un verre fixé à un bâton, ainsi que de divers râteaux, « chats », dragues, bennes de fond, silos, etc.
Collecte de périphyton. Afin d'étudier la composition spécifique du périphyton, la plaque à la surface de divers objets sous-marins (cailloux, graviers, pierres, tiges et feuilles de plantes supérieures, coquilles de mollusques, parties en bois et en béton d'ouvrages hydrauliques, etc.) est éliminée à l'aide d'un couteau ordinaire ou grattoirs spéciaux. Cependant, de nombreux organismes intéressants meurent au cours du processus ; certains d'entre eux sont emportés par les courants d'eau, les organes de fixation des algues au substrat sont détruits, l'image du placement mutuel des composants de la biocénose est perturbée. Par conséquent, il est préférable de collecter les algues avec le substrat, qui est complètement ou partiellement soigneusement retiré à la surface de l'eau afin que le courant n'en emporte pas les algues. Le substrat extrait (ou son fragment), avec les algues, est placé dans un récipient préparé pour l'échantillon et rempli avec seulement une petite quantité d'eau du même réservoir afin d'étudier plus avant le matériel collecté à l'état vivant ou avec un Solution de formaldéhyde à 4 %. Les algues terrestres ou aériennes sont collectées, si possible, avec le substrat dans des sacs en papier stériles ou dans des récipients en verre avec une solution de formaldéhyde à 4 %.
Étiquetage et fixation des échantillons. Tenir un journal de terrain. Pour étudier les algues à l'état vivant et fixe, le matériel collecté est divisé en deux parties. Le matériel vivant est placé dans des récipients en verre stériles (tubes à essai, flacons, bocaux), fermés par des bouchons de coton et non remplis à ras bord, ou dans des sacs en papier stériles. Afin de mieux maintenir les algues en vie dans des conditions expéditionnaires, des échantillons d'eau sont emballés dans du papier d'emballage humide et placés dans des boîtes. Les échantillons doivent être périodiquement déballés et exposés à une lumière diffuse pour soutenir les processus photosynthétiques et enrichir l'environnement en oxygène.
Les échantillons collectés sont soigneusement étiquetés. Les étiquettes, remplies d'un simple crayon ou de pâte, indiquent le numéro de l'échantillon, l'heure et le lieu de collecte, l'outil de collecte et le nom du collecteur. Les mêmes données sont consignées dans le journal de terrain, dans lequel, en plus, les résultats des mesures de pH, de température de l'eau et de l'air, un dessin schématique, une description détaillée du réservoir étudié, la végétation aquatique supérieure qui s'y développe, et d'autres les observations sont saisies.
Etude qualitative du matériel collecté. Le matériel est préalablement examiné au microscope à l'état vivant le jour de la collecte afin de constater l'état qualitatif de l'algue avant l'apparition des modifications provoquées par le stockage du matériel vivant ou la fixation des échantillons (formation de cellules reproductrices, colonies , perte de flagelles et de motilité, etc.). A l'avenir, il est étudié en parallèle à l'état vivant et figé. Des préparations sont préparées pour l'examen microscopique des algues: une goutte du liquide à l'étude est appliquée sur une lame de verre et recouverte d'une lamelle. Si les algues vivent hors de l'eau, elles sont placées dans une goutte d'eau du robinet ou de glycérine hydratée. Si des observations à long terme du même objet sont nécessaires, la méthode de la goutte suspendue donne un bon résultat. Une petite goutte du liquide de test est appliquée sur une lamelle propre, après quoi la lamelle, dont les bords sont enduits de paraffine, d'huile de paraffine ou de gelée de pétrole, est placée sur une lame de verre spéciale avec un trou dans le milieu pour que la goutte ne touche pas le fond du trou. Une telle préparation peut être étudiée pendant plusieurs mois, en la conservant entre les travaux en chambre humide. Lors de l'identification des algues, il est nécessaire d'atteindre la précision de leur détermination. Lors de l'étude du matériel d'origine, il est nécessaire de noter tout écart, même mineur, de taille, de forme et d'autres caractéristiques morphologiques, de les corriger dans des descriptions, des dessins et des microphotographies.
Méthode de comptabilisation quantitative des algues. Des échantillons de phytoplancton, de phytobenthos et de périphyton peuvent faire l'objet d'une comptabilité quantitative. Les données d'abondance des algues sont les données initiales pour déterminer leur biomasse et recalculer d'autres indicateurs quantitatifs par cellule ou unité de biomasse. Le nombre d'algues peut être exprimé en nombre de cellules, coenobia, segments de fils d'une certaine longueur, etc. Le nombre d'algues planctoniques est compté à l'aide de chambres de comptage (Fuchs-Rosenthal, Nazhott, Goryaev, etc.) avec un microscope grossissement de 420 fois. La quantité moyenne d'algues obtenue à partir d'au moins trois comptages est recalculée pour un volume d'eau donné. Étant donné que le substrat pour le règlement des algues peut être des objets sous-marins (pierres, pieux, plantes, animaux, etc.), dans certains cas, la quantité d'algues est calculée par unité de surface, dans d'autres - par unité de masse.
1.3 POSITION DES ALGUES DANS LE SYSTEME ACTUEL DU MONDE ORGANIQUE
Pour créer un système naturel du monde organique, les taxonomistes utilisent un ensemble des caractéristiques les plus significatives des organismes inclus dans une catégorie taxonomique particulière. Ces signes comprennent :
1) le développement historique d'un groupe d'organismes vivants à partir de restes fossiles ;
2) caractéristiques de la structure morphologique et anatomique des espèces modernes ;
3) caractéristiques de la reproduction et du développement embryonnaire ;
4) caractéristiques physiologiques et biochimiques ;
5) caryotype, déterminé par le nombre, la taille et la forme des chromosomes ;
6) type de nutriments de réserve
7) distribution sur notre planète et un certain nombre d'autres.
Le système généralement accepté du monde organique n'a pas encore été créé. Jusqu'à présent, le nombre d'empires, de royaumes, de sous-royaumes, de types (départements) distingués par différents auteurs n'est pas le même. Un moment fondamentalement nouveau dans ce système du monde organique par rapport au précédent est l'attribution du royaume de Protista. Le nom "royaume de Protista" ( protiste) proposé en 1866 par E. Haeckel. Pendant la majeure partie du 20e siècle, les partisans de la séparation des protistes en un royaume séparé ont renforcé leur position, bien qu'ils en aient exclu les bactéries et les éponges, mais l'ont complétée avec le reste des protozoaires, ainsi que certains champignons et algues. Fait actuellement partie du royaume protiste de nombreux auteurs incluent tous les organismes eucaryotes unicellulaires et coloniaux, quel que soit le type de nutrition et de fonctionnement. Cela signifie qu'ils sont considérés comme un niveau particulier d'organisation de la matière vivante. Comprendre les protistes précisément comme prétissus (plutôt qu'unicellulaires) permet à divers auteurs de systèmes d'inclure dans leur composition (selon ce que l'auteur entend par tissu) tout ou partie des groupes d'algues multicellulaires (vertes, rouges, brunes), des organismes ressemblant à des champignons, ou "pseudo-champignons" - hypochytridies ( Hyphochytridiomycota), les oomycètes ( Oomycète) et labyrinthique ( Labyrinthulomycota). En conséquence, le royaume de Protista a uni un groupe d'organismes extrêmement hétérogènes, dont certains étaient auparavant inclus dans le royaume des animaux (protozoaires), le royaume des champignons (acrasia et mixomycètes plasmodiaux, la plupart des champignons inférieurs - chytridiomycètes et oomycètes ), ainsi que dans le règne végétal (euglènes, dinophytes, cryptophytes. , diatomées, algues dorées, jaune-vert, vertes).
Ainsi, la taxonomie moderne des algues se caractérise par la présence de nombreux systèmes qui diffèrent plus ou moins les uns des autres non seulement au niveau des petits taxons (genres, familles, ordres, classes) mais aussi aux niveaux taxonomiques les plus élevés. (départements, sous-royaumes, royaumes,) . Par exemple, les charophytes d'un même volume sont considérées par différents auteurs comme un département, une classe, voire un ordre. De plus, dans un système, ils sont affectés au royaume des plantes, dans un autre - au royaume protiste ou Chromiste. Parallèlement, selon un certain nombre de caractéristiques essentielles (présence, comme dans les plantes vertes, de chlorophylle un, caroténoïdes, ainsi que les phycobilines comme dans les algues rouges, type de photosynthèse oxygénée, etc.) les cyanobactéries sont très similaires aux algues. À cet égard, elles sont souvent appelées algues bleues et sont considérées dans le cours d'algologie.
2. CARACTERISTIQUES DES ALGUES
2.1 DIVISION JAUNE VERT ( XANTHOPHYTE)
Le département des algues jaune-vert comprend des organismes qui se trouvent à différents stades de différenciation morphologique du thalle - unicellulaire, colonial et multicellulaire. Parmi eux, on trouve principalement des structures coccoïdes, palmelloides ou filamenteuses, moins souvent - amiboïdes, monadiques, siphonales et lamellaires. Les formes mobiles d'algues jaune-vert (y compris les zoospores) se caractérisent par la présence de deux flagelles de taille inégale (latéral - court, en forme de coléoptère et antérieur - long avec mastigonèmes) et la couleur jaune-vert des chromatophores, due à la présence de chlorophylles a et c, de carotènes in et e, de xanthophylles (anthéraxanthine, lutéine, zéaxanthine, washriaxanthine, violaxanthine et néoxanthine). Selon la prédominance de certains pigments, il existe des espèces de couleur jaune clair ou foncé, moins souvent vertes et dans certaines bleues. Produits de rechange - volutine, graisse, souvent chrysolaminarine. Dans les formes primitives, le contenu de la cellule est entouré d'un mince périplaste, tandis que chez les représentants plus organisés, il existe une membrane de pectine ou de cellulose (solide ou bicuspide). La membrane cellulaire est souvent imprégnée de sels de fer, de silice, de chaux et présente diverses décorations sculpturales.
Il existe plusieurs chromatophores dans le protoplaste de la cellule, qui peuvent être en forme de disque, lamellaire, en forme de ruban ou de coupe ou en forme d'étoile. Un noyau ou plusieurs. Certaines espèces ont des pyrénoïdes. Les formes mobiles ont un stigmate. Les algues jaune-vert peuvent se reproduire par division cellulaire longitudinale, désintégration de colonies ou de filaments en sections séparées, ainsi que par des zoospores ou des aplanospores. Le processus sexuel (iso- ou oogamie) est connu de quelques-uns. Pour supporter des conditions défavorables, certaines espèces forment des kystes avec une membrane bivalve faiblement silicifiée. Les algues de cette division se trouvent principalement dans les réservoirs d'eau douce propres, moins souvent dans les mers et les eaux et sols saumâtres.
2.1.1 Classe Xanthophytacées ( Xanthophycées)
Cette classe comprend les organismes unicellulaires et multicellulaires, principalement de structure coccoïde ; moins fréquemment, une structure corporelle monadique, rhizopodiale, palmelloide, filamenteuse, multifilamenteuse ou siphon est observée. Formes et stades monadiques avec deux flagelles inégaux et un stigmate situé sur en pointe chromatophore, sous sa coquille. Les chromatophores sont entourés d'un canal du réticulum endoplasmique qui se prolonge dans la membrane externe de l'enveloppe nucléaire.
Conformément aux types d'organisation du thalle, la classe est divisée en six ordres: hétérochlorure ( Hétérochloridales), rhizochlorure ( Rhizochloridales), hétérogle ( hétérogloéales), mischococque ( mischococoques), botridien ( Botrydiales) et tribônémique ( Tribonematales).
Commander Botridium ( Botridiales). Les représentants de l'ordre sont caractérisés par une structure siphonale du thalle. Extérieurement, ils peuvent avoir une forme complexe, mais selon la structure du protoplaste, ils constituent une cellule multinucléée géante. En règle générale, le thalle se différencie en parties souterraines colorées et incolores. Les représentants typiques de l'ordre sont les genres Botridium et Vosheria.
Genre botridium ( Botrydium) associe des algues siphons fixées terrestres ayant une forme sphérique, en forme de poire ou de massue. Leur partie souterraine est un système dichotomique ramifié de rhizoïdes incolores. La cellule est recouverte d'une membrane de pectine qui, saturée de chaux, grossit avec l'âge. Des chromatophores lamellaires ou en forme de disque et de nombreuses gouttes d'huile sont situés dans la couche de paroi du cytoplasme. Les petits noyaux ne sont visibles qu'après coloration.
Botridium se reproduit par zoospores, parfois par autospores ou aplanospores. Dans des conditions défavorables (séchage prolongé), le contenu de la partie terrestre (boule) se déplace dans les rhizoïdes et se décompose en parties séparées recouvertes d'une coquille épaisse, formant des kystes au repos - rhizocystes. Avec l'apparition de conditions favorables, les rhizocystes germent en de nouveaux individus directement ou via le stade zoospore.
Plus de 10 espèces sont connues, dont 1 espèce - B. granulaire ( B. granulat) - en Biélorussie. Ils se développent sur des dépôts argilo-limoneux au bord des plans d'eau, au fond des étangs d'assèchement, dans les ornières des routes de campagne et forestières, sur des sols humides, riches en éléments nutritifs et à forte teneur en chaux.
Clan Vosheria ( Vaucheria) comprend des algues dont le thalle est constitué de filaments irrégulièrement et rarement ramifiés de délicate couleur vert clair avec des rhizoïdes ramifiés incolores. Il s'agit d'une cellule multinucléée géante. Sa partie centrale est occupée par une grande vacuole à sève cellulaire. De nombreux chromatophores en forme de disque sans pyrénoïdes ni gouttelettes d'huile sont situés dans la couche de paroi du cytoplasme.
La reproduction asexuée est réalisée par des zoospores et aplanospores multiflagellées et multinucléaires. Dans le même temps, le contenu aux extrémités des branches devient plus dense et plus sombre, est séparé par un septum du fil commun et se transforme en zoosporange, où se forme une grande zoospore avec de nombreux flagelles appariés le long de la périphérie.
Le processus sexuel chez Vosheria est oogame. L'anthéridie et une ou deux ou plusieurs oogones sont formées sur un fil ou sur des branches courtes spéciales. Lorsque l'œuf mûrit, une goutte de contenu émerge du bec de l'oogone, attirant les spermatozoïdes. L'un d'eux (avec deux flagelles de longueur inégale) est introduit dans l'oogone par le trou formé et féconde l'œuf. Après la fécondation, une oospore se développe dans l'oogonium avec une coquille épaisse contenant beaucoup d'huile et d'hématochrome. Après une période de dormance, la division de réduction du noyau s'y produit et il se transforme en un nouveau fil haploïde.
62 espèces sont connues, réparties dans le monde entier. En Biélorussie, 1 espèce est notée - Vaucheria De Candolle sp.
Ordre tribonemal ( Tribonematales). Il associe des formes caractérisées par une structure filamenteuse du thalle. Ce sont les représentants les plus organisés des algues jaune-vert. En apparence, ils ressemblent aux ulothrix du département des Verts et à de nombreuses espèces du département des Algues dorées. Un représentant typique de cet ordre est le genre Triboneme.
Genre Tribonema ( Tribonema) comprend les algues dont les filaments ne sont pas ramifiés. Au début, ils sont attachés à un substrat à l'aide d'une cellule basale, puis, en raison de sa mort, ils flottent à la surface du réservoir et se retrouvent déjà flottant librement, formant une boue jaune-vert. Un trait caractéristique par lequel les filaments de tribonema se distinguent facilement des autres algues filamenteuses est le contour particulier de leurs extrémités en forme de deux cornes. Cela est dû au fait que la membrane cellulaire du tribonème est forte, bicuspide et se compose de deux moitiés identiques. Le bord d'une moitié est sur le bord de l'autre au milieu de la cage. Au cours de la division cellulaire, une section cylindrique d'une nouvelle membrane est formée à partir de sa partie médiane, dans laquelle un septum transversal est posé. Ainsi, les moitiés des cellules voisines sont fermement connectées les unes aux autres, et lorsque le fil se brise en morceaux ou se brise en cellules individuelles, des fragments caractéristiques en forme de H de la coquille se forment. Il y a généralement plusieurs chromatophores dans chaque cellule du tribonème, en forme de disque, sans pyrénoïdes. Lors de la reproduction, une ou deux zoospores multiflagellées ou aplanospores se forment dans les cellules, à la sortie desquelles les valves divergent et le fil d'algue se désagrège. Pour supporter des conditions défavorables, des akinètes à paroi cellulaire épaisse ou à kystes sont utilisés. 22 espèces de tribonemes sont connues, dont 6 en Biélorussie. Distribué principalement dans la zone côtière de divers réservoirs sur des plantes aquatiques, des pierres, certaines - dans le sol; forment souvent des mottes molles ressemblant à du coton, non mucilagineuses, jaune-vert.
2.2 DIVISION ALGUES BRUNES ( PHÉOPHYTES)
Le département Algues brunes comprend de nombreuses algues multicellulaires, principalement macroscopiques, de structure simple et complexe. Leurs tailles varient de quelques millimètres à plusieurs mètres (parfois jusqu'à 60 m ou plus). Le thalle se développe à la suite d'une croissance intercalaire ou en raison de l'activité de la cellule apicale. En apparence, ce sont des buissons ramifiés, des croûtes, des plaques, des cordons, des rubans, disséqués de manière complexe en organes en forme de tige et de feuille. Les thalles de certains grands représentants ont des bulles d'air qui maintiennent les branches dans l'eau en position verticale. Pour la fixation au sol, on utilise des rhizoïdes ou des excroissances en forme de disque à la base du thalle - le disque basal.
Selon la différenciation morphologique et anatomique du thalle, les algues brunes se situent à un niveau supérieur à tous les autres groupes. Parmi eux, on ne connaît ni les formes unicellulaires ni coloniales, ni les thalles en forme de simple fil non ramifié. Le thalle des algues brunes vivantes les plus simples est hétérotriche, alors que la grande majorité possède des thalles d'une structure tissulaire fausse ou vraie (on distingue les tissus assimilateurs, de stockage, mécaniques, conducteurs).
La coquille des cellules est mucilagineuse à l'extérieur, se compose de substances pectines et d'une couche interne de cellulose. Le mucus protège les cellules des influences mécaniques, du dessèchement à marée basse, etc. Dans le cytoplasme, il y a un noyau et des chromatophores d'un disque, moins souvent de forme rubanée ou lamellaire, des vacuoles, dans de nombreux types de pyrénoïdes.
Les chromatophores des cellules d'algues brunes contiennent des chlorophylles a et c, des carotènes et plusieurs xanthophylles - fucoxanthine, violaxanthine, antheraxanthine et zéaxanthine. Ces pigments déterminent la couleur brune des algues. Les produits de base sont le polysaccharide laminarine, l'alcool hexahydrique mannitol et les lipides. Chez les algues brunes, les deux formes de reproduction se produisent : asexuée et sexuée.
La reproduction asexuée est réalisée par des sections du thalle. Certaines algues ont des brindilles spécialisées (bourgeons de couvain) qui se détachent facilement et produisent de nouvelles plantes. De plus, dans la plupart des algues brunes, la reproduction asexuée se produit par le biais de zoospores, chez certains représentants - les tétraspores et chez une seule espèce - les monospores. Les zoospores se développent dans des sporanges unicellulaires ou multicellulaires. La formation des spores est précédée de la méiose (à l'exception des cyclospores, dans lesquelles la méiose se produit avant la formation des gamètes).
Le processus sexuel est iso-, hétéro- et oogame. Avec l'iso- ou l'hétérogamie, les gamètes se forment dans des gamétanges multicellulaires et multichambres, qui peuvent se développer à partir d'une ou de plusieurs cellules. Chez les algues brunes les plus organisées, le processus sexuel est oogame. L'œuf est fécondé à l'extérieur de l'oogone. Le zygote sans période de dormance germe en une plante diploïde.
Pour la plupart des algues brunes, une modification des formes de développement est caractéristique : pour certaines elle est isomorphe, pour d'autres elle est hétéromorphe. Ces différents types de cycle de vie servaient auparavant de base pour diviser les Algues Brunes en 3 classes : isogénérées avec un cycle de développement isomorphe, hétérogénérées avec un cycle de développement hétéromorphe, et cyclosporiques avec un ordre Fucal, où il n'y a pas d'alternance de générations. Cependant, la division des algues brunes en isogénères et hétérogènes est plutôt arbitraire, car dans les deux classes, il existe des représentants avec le type opposé de changement dans les formes de développement. Par conséquent, une approche plus correcte de la classification des algues brunes consiste à les diviser en deux classes - Phaeozoospores ( Phéozsurosporophycées) et Cyclosporacées ( Cyclosporophycées).
Presque toutes les algues brunes vivent dans les mers en tant qu'organismes benthiques, épiphytes ou planctoniques secondaires. Les fourrés d'algues brunes sont des lieux de nourriture, de reproduction et d'abri pour de nombreuses espèces animales, un substrat pour les micro- et macro-organismes et l'une des principales sources de matière organique aux latitudes tempérées et subpolaires. Ils sont largement utilisés dans l'industrie (alimentaire, parfumerie, textile) en raison de la présence de substances précieuses telles que l'acide alginique, l'alginate, le mannitol, etc.
2.2.1 Classe Phaeozoosporophyceous ( Phaeozosursporophycées)
La plupart des algues de la classe Phaeozoosporophyceous sont caractérisées par 2 formes de développement indépendantes - sporophyte et gamétophyte ou gamétosporophyte, qui peuvent être similaires en apparence, structure et taille et différentes, c'est-à-dire il y a un changement isomorphe et hétéromorphe dans les formes de développement. Chez les représentants primitifs, il n'y a pas de changement dans les formes de développement.
La classe Phaeozoosporophyceous est divisée en 11 ordres, dont 5 sont donnés ci-dessous.
Commander Ectocarnal ( Ectocarpiens). Comprend les algues brunes, les thalles (à la fois sporophytes et gamétophytes) qui sont construits à partir de filaments à une seule rangée capables de se ramifier. Leurs tailles varient de microscopiques à 30 centimètres et plus. Ces algues forment des plaques ou des buissons sur des rochers ou d'autres algues. Ils se reproduisent de manière asexuée et sexuée. Les organes reproducteurs sont des réceptacles simples et multiples. Les uniloculaires sont toujours des sporanges, tandis que les multiloculaires peuvent également fonctionner comme des gamétanges.
Espèce du genre Ectocarpus (ectocarpe) ont un thalle touffu de 0,1 à 30 cm de haut constitué de minces filets verticaux rampants et ramifiés à une rangée. La croissance des filaments est intercalaire ou diffuse. La fixation au substrat est réalisée par des rhizoïdes qui, chez les gros spécimens, forment une sorte d'écorce à la base des branches. Au sommet des branches, les cellules se rétrécissent et se terminent par un long poil incolore.
Les sporanges et les gamétanges sont disposés en excroissances latérales des branches. À l'intérieur des sporanges uniloculaires, la méiose et la mitose se produisent, suivies de la formation de zoospores biflagellées. Les zoospores se transforment en organismes dioïques haploïdes avec des gamétanges multicellulaires. Les gamètes sont isomorphes, mais diffèrent par leur comportement : la femelle perd sa mobilité et libère une substance odorante qui attire les gamètes mâles, dont l'un la féconde. Le zygote sans période de dormance germe en un sporophyte diploïde.
Ordre sfacelarial ( Sphacelariales). Comprend des algues aux thalles durs et buissonnants de quelques millimètres à 30 cm de haut; branches cylindriques. Contrairement aux autres algues brunes, chez les algues sphacéliales, chaque branche se termine par une grande cellule, en raison de la division de laquelle les algues ont une croissance strictement apicale. Leur thalle est caractérisé par une base en forme de plaque corticale de plusieurs couches de cellules.
La reproduction végétative se fait par l'intermédiaire de stolons (filaments de plusieurs rangées de cellules rampant sur le sol) ou de bourgeons de couvain spéciaux qui se séparent des branches. Les sphacélaires ont un changement isomorphe dans les formes de développement.
Algue genre sphacelaria (Sphacelaria) trouve dans toutes les mers. Le thalle de ses représentants a l'apparence d'un buisson atteignant 4 cm de haut, constitué d'une semelle lamellaire et de fils ramifiés s'étendant à partir de celle-ci. Chaque ramification du fil au sommet porte une grande cellule, qui ne se divise que dans le sens transversal et provoque la croissance du thalle en longueur. Les cellules détachées de cette manière sont ensuite divisées dans le sens longitudinal, grâce à quoi des cellules étroites se forment, et le thalle devient multicouche et constitué vers l'extérieur, pour ainsi dire, de segments.
Commande coutellerie ( Coutellières). Il comprend les algues brunes, qui se caractérisent par une structure trichothallique du thalle due à la zone de croissance située dans la partie basale des poils multicellulaires, qui sont situés sur les bords du thalle lamellaire ou au sommet des branches du thalle touffu. Les cellules de la zone de croissance se divisent, séparant les cellules vers la périphérie et vers le thalle.
Ordre dictatorial ( Dictyotes). Combine des espèces caractérisées par une croissance apicale et une ramification généralement dichotomique dans le même plan. Reproduction asexuée au moyen d'aplanospores (tétraspores). Le processus sexuel est oogame. Le changement des formes de développement est isomorphe. La plupart des dictyotales poussent dans les mers tropicales et subtropicales. On les trouve assez souvent dans les mers Noires (espèces des genres dictyota, dilofus et padina) et dans la mer du Japon (dictyota).
Sortes sorte de dictée (Dictyota) sont caractérisés par un thalle ramifié fourchu à plat, généralement situé dans le même plan des branches sans nervure longitudinale. Le thalle se développe à partir d'un rhizome cylindrique attaché au substrat par des rhizoïdes. Le sommet de chaque branche se termine par une grande cellule . À l'intérieur des branches, il y a une couche de grandes cellules incolores, entourées à l'extérieur par une écorce d'une couche de petites cellules intensément colorées.
Sur les sporophytes, les sores des sporanges unicellulaires se développent à partir des cellules de surface, où se forment quatre tétraspores immobiles. Les tétraspores germent en gamétophytes. Dictyota est une algue dioïque : sur les gamétophytes femelles, des sores d'ovogonies à un seul nid se forment avec un œuf chacun. Les anthéridies sont produites sur les gamétophytes mâles. Les œufs sont libérés de l'oogone et sont fécondés par des spermatozoïdes dans l'eau. Le zygote germe immédiatement dans un nouvel organisme - le sporophyte. Le dictyota le plus courant est dichotomique (ré. dichotome).
Genre varech (Laminaire) comprend les espèces dont le thalle est divisé en une plaque en forme de feuille, un tronc et des rhizoïdes . Les plaques foliacées sont régulières ou ridées, entières ou disséquées. Le tronc et les rhizoïdes sont vivaces, la plaque en forme de feuille change chaque année. Sur les coupes longitudinales du pétiole et des organes de fixation, leur structure anatomique assez complexe est révélée. La partie externe du pétiole est un cortex composé de plusieurs couches de cellules avec des chromatophores ; la couche intermédiaire est représentée par un grand tissu de stockage cellulaire et, enfin, l'intérieur (noyau) - conducteur et mécanique. Le système conducteur comprend des filaments tubulaires avec des extensions en forme d'entonnoir au niveau des parois cellulaires. Ces cloisons ont des pores et sont appelées plaques de tamis, et les fils - tubes tamis . Le pétiole croît en épaisseur en raison de la division des cellules corticales, qui se produit périodiquement, à la suite de quoi des couches concentriques sont clairement visibles sur la section transversale du pétiole, ressemblant aux anneaux de croissance des plantes supérieures.
Lors de la reproduction à la surface de la plaque en forme de feuille de cellules corticales, des groupes (sori) forment des zoosporanges uniloculaires, chacun formant de 16 à 128 zoospores à deux flagellations. Dans des conditions favorables, les zoospores germent en excroissances filamenteuses microscopiques - gamétophytes mâles et femelles.
Le processus sexuel du varech est oogame. L'œuf mature quitte l'oogone et se fixe à son extrémité supérieure. Dans cette position, la fécondation se produit. Le zygote se développe en sporophyte sans période de dormance. Le gamétophyte femelle assure non seulement la formation des cellules germinales, mais également le lieu de fixation du futur sporophyte.
2.2.2 Classe Cyclosporophycea ( DEyclosporophycées)
La classe Cyclosporophyceous comprend les algues qui n'ont pas d'alternance de générations dans le cycle de développement. Leurs thalles diploïdes ne portent que des organes de reproduction sexuée, qui se développent dans des réceptacles arrondis spéciaux - les conceptacles ou scaphidia. La méiose chez les cyclosporides se produit avant la formation des gamètes. Il n'y a pas de reproduction asexuée par les spores. Tous les cyclosporophyces sont de grandes algues. Ordre de Fucalie (Fucales). Combine les algues, qui se caractérisent par une forme touffue de thalle à croissance apicale. Les cellules des parties axiales du thalle sont faiblement divisées. Ils sont allongés et forment le noyau. genre fucus (Fucus) comprend des espèces avec un thalle plat, en forme de ceinture, ramifié de manière dichotomique jusqu'à 1 m de long.Une veine médiane court le long des lobes du thalle avec des bords lisses ou dentelés, passant dans la partie inférieure dans un pétiole, qui est attaché au substrat par un disque basal. Dans certains types de fucus, des bulles d'air sous forme de gonflements sont situées sur les côtés de la nervure médiane. Le thalle se développe en raison de l'activité des cellules apicales. Lors de la reproduction, les extrémités du thalle gonflent, prennent une couleur jaune-orange clair et se transforment en réceptacles sur lesquels se forment des scaphidies trouées. Entre les paraphyses sur les parois du scaphidium femelle, des ovogonies se forment, tandis que le mâle - des anthéridies. Le zygote germe sans période de dormance. Les espèces de Fucus sont communes le long des côtes des mers froides et tempérées de l'hémisphère nord, formant souvent de grands fourrés dans la zone intertidale, ce qui facilite leur collecte et leur utilisation. Les espèces de Fucus sont utilisées comme engrais, aliments pour le bétail, farine alimentaire, alginates et autres produits chimiques. Dans les mers de Russie, il existe 5 espèces de ce genre. Le plus célèbre F. vesiculosus (F. vésiculeux) et F. bilatérale (F. distique).
2.3 Département Algue Rouge, ou Pourpre (RHODOPHYTA)
L'écrasante majorité des représentants du département sont des organismes multicellulaires de structure morphologique et anatomique complexe, et seuls quelques-uns, les plus primitifs, ont un thalle unicellulaire ou colonial de structure coccoïde. De nombreuses algues violettes sont de grandes algues, atteignant une longueur de plusieurs centimètres à deux mètres, mais parmi elles, il existe de nombreuses formes microscopiques.
Dans la forme, les algues rouges se présentent sous forme de filaments, de buissons, de plaques, de bulles, de croûtes, de coraux, etc. Les formes lamellaires atteignent une grande variété. Il y a des plaques entières et disséquées de manière complexe, avec des excroissances supplémentaires le long du bord et à la surface. Certains pourpres sont fortement calcifiés et ressemblent à des fossiles.
Avec toute la variété des formes externes, les algues rouges se caractérisent par un plan structurel unique du thalle - il est basé sur une structure hétérotriche chez toutes les algues violettes multicellulaires.
Les branches d'algues rouges se divisent en deux catégories. Certaines sont les principales branches longues qui poussent en longueur pendant toute la période de croissance des plantes, les soi-disant branches à croissance illimitée. D'autres ne poussent que jusqu'à une certaine limite et restent toujours plus ou moins courts - ce sont des branches à croissance limitée. De plus, ils ont également des branches spécialisées qui agissent comme des antennes ou des rhizoïdes, qui servent à une fixation ou une adhérence supplémentaire les unes aux autres. Le type d'organisation parenchymateuse est pratiquement absent. Le seul exemple d'un tel thalle est un représentant de la classe des Bangiacées (porphyre). Dans la majorité des thalles violets, le type pseudoparenchymateux (dû à l'entrelacement des branches d'un axe - une structure uniaxiale ou plusieurs - multiaxiale). Une augmentation de la taille des thalles dans les formes primitives est réalisée en raison de la division cellulaire diffuse, dans les plus organisées, à la suite de la division des cellules apicales, et dans un certain nombre d'espèces, en raison du méristème apical ou marginal. Les organes de fixation au substrat sont les rhizoïdes, les ventouses, les soles ou les plaques rhizoïdales rampantes.
Les cellules des algues rouges sont recouvertes d'une membrane dans laquelle se distinguent la couche interne, cellulosique, et externe, pectine, mucilagineuse. L'agar-agar obtenu à partir de ce dernier contient, en plus de la pectine, des sucres et des protéines. L'enveloppe peut être imprégnée de sels de chaux, de magnésium ou de fer. Le cytoplasme se distingue par une viscosité accrue, adhère étroitement aux parois et est sensible aux changements de salinité du milieu. Dans les algues hautement organisées, les cellules sont multinucléées, dans les moins organisées, elles sont mononucléaires.
La forme des chromatophores dépend de l'intensité de l'éclairement, de la taille et de l'âge des cellules. Cependant, plus l'organisation de l'algue est élevée, plus il y a de chromatophores dans ses cellules et plus leur forme (principalement lenticulaire) est constante. Les pyrénoïdes sont absents chez de nombreuses espèces. Comme les autres algues, la couleur des plastes et de tout le corps des algues rouges est due à une combinaison de plusieurs pigments : les chlorophylles a et d, les phycobilines (phycocyanine, phycoérythrine, allophycocyanine) et les caroténoïdes. La couleur du thalle varie du rouge cramoisi (prédominance de la phycoérythrine) au bleu-acier (avec un excès de phycocyanine). Les méthodes de reproduction des algues rouges sont très diverses. La reproduction végétative n'est propre qu'aux primitives. Elle est réalisée en raison de la formation de pousses supplémentaires, de la croissance d'un nouveau thalle à partir de la semelle de l'ancien, mort, et également par division cellulaire. Des sections arrachées de thalles meurent. La reproduction asexuée est réalisée par des mono-, bi-, tétra- et polyspores formées dans les sporanges. Les tétraspores se forment sur des plantes asexuées diploïdes - sporophytes (tétrasporophytes). Dans les tétrasporanges, la méiose se produit avant la formation des tétraspores.
Le processus sexuel est oogame. Karpogon se compose généralement d'une partie basale élargie - l'abdomen (avec un noyau à l'intérieur) et d'une excroissance tubulaire - le trichogyne, qui reçoit le sperme. Les spermatanges sont de petites cellules incolores dont le contenu est libéré sous la forme de petits gamètes mâles nus et dépourvus de flagelles - spermatozoïdes.
La fécondation de l'ovule est réalisée en déplaçant le sperme le long du trichogyne dans le carpogon. Après la fécondation, la partie basale du carpogon est séparée par un septum du trichogyne, qui meurt et subit un développement ultérieur, conduisant à la formation de carpospores. Les détails de ce développement sont importants dans la classification des violets. Dans certaines algues rouges, le contenu du zygote est divisé avec la formation de spores nues immobiles - carpospores, dans d'autres, un système de filaments spéciaux est formé à partir du carpogon fécondé - gonimoblastes, dont les cellules se transforment en carposporanges, produisant un carpospore chaque. Dans la plupart des plantes violettes, le développement des carpospores a lieu avec la participation de cellules auxiliaires. Dans de tels cas, le gonimoblaste ne se développe pas à partir de l'abdomen du carpogon, mais à partir de la cellule auxiliaire. Si les cellules auxiliaires sont retirées du carpogon, des fils de connexion (oblastémiques) se développent à partir de son abdomen après la fécondation; leurs cellules sont diploïdes. Les filaments oblastémiques se développent jusqu'aux cellules auxiliaires et se dissolvent au point de contact, après quoi la plasmogamie se produit, entraînant le développement d'un gonimoblaste avec des carpospores - un carposporophyte. Par conséquent, les cellules auxiliaires remplissent une fonction auxiliaire - elles stimulent la division du noyau cellulaire du fil conjonctif et fournissent une nutrition supplémentaire. Chez les algues rouges les plus organisées (florideeficiaceae), des cellules auxiliaires se développent après fécondation du carpogon dans son voisinage immédiat. Les filaments ooblastiques ne se forment pas dans ces algues. La cellule auxiliaire, située à côté de l'abdomen du carpogon, se confond avec lui et forme un procarpe.
Les cycles de développement des algues rouges sont variés. Chez certains représentants des florideeficiaceae, il y a un changement dans trois formes de développement: gamétophyte haploïde, carpo- et tétrasporophyte diploïde. Dans ce cas, le zygote se divise sans réduction du nombre de chromosomes, formant un sporophyte, sur lequel, à la suite de la méiose, des tétraspores se forment, donnant naissance à des gamétophytes. Ainsi, il existe deux formes libres de la même plante - le tétrasporophyte et le gamétophyte. Chez d'autres algues (avec un changement hétéromorphe des formes de développement), le tétra- et le carposporophyte sont souvent peu développés et même réduits, parfois le gamétophyte est réduit (il se forme sur le sporophyte).
...Documents similaires
L'étude des types et des caractéristiques des algues - des organismes primitifs qui n'ont pas d'organes, de tissus et de vaisseaux sanguins complexes. Revue des principaux processus physiologiques des algues : croissance, reproduction, nutrition. Classification et évolution des algues du sol et de l'eau.
résumé, ajouté le 06/07/2010
Caractéristiques biologiques des algues, leur structure anatomique. la reproduction algues unicellulaires. Orientations pour le développement de l'algologie appliquée. Origine et évolution des algues, leurs groupes écologiques. Algues habitats aquatiques, neige, glace.
présentation, ajouté le 25/11/2011
Les algues en tant que représentants des organismes photoautotrophes de notre planète, leur origine et leurs stades de développement. Modes et conditions de nutrition des algues. Reproduction de leur propre espèce dans les algues par reproduction végétative, asexuée et sexuée.
résumé, ajouté le 18/03/2014
Caractéristiques générales des algues vertes - un groupe de plantes inférieures. Habitat des algues vertes marines. Leur reproduction, structure et méthodes de nutrition, composition chimique. Description des types d'algues les plus courants dans la mer du Japon.
résumé, ajouté le 16/02/2012
La division des algues en groupes systématiques du rang le plus élevé, sa coïncidence avec la nature de la couleur et les caractéristiques structurelles. Parois cellulaires des algues. Reproduction asexuée et sexuée des algues. Similitudes et différences entre les algues jaune-vert et vertes.
résumé, ajouté le 06/09/2011
Département des algues bleues (Cyanophyta). Classes et types d'algues. Structure cellulaire. Protoplaste. Composants du protoplaste. Plasmodesme. Pseudovacuoles. Résistance des algues bleues aux effets d'un assombrissement prolongé. La reproduction.
conférence, ajouté le 01/06/2008
Méthodes de nutrition et structure cellulaire des algues. Les principaux types de structure morphologique de leur corps. Analyse comparative de la diversité spécifique de diverses espèces d'algues dans les écotopes. Reproduction, cycles de développement et distribution des plantes dans les plans d'eau.
dissertation, ajouté le 12/05/2014
Méthodes de nutrition et principaux types de structure morphologique du corps des algues. La structure de leurs cellules, les cycles de reproduction et de développement. Analyse comparative de la diversité spécifique de diverses espèces d'algues dans les écotopes. Collecte de matériel et herboristerie de plantes.
dissertation, ajouté le 12/11/2014
Chaîne trophique de l'écosystème du réservoir. Classification des algues, leur distribution en fonction de la profondeur, répartition et rôle dans les biogéocénoses. Utilisation humaine des algues. Reproduction végétative, asexuée, sexuée. Groupes d'algues du sol.
présentation, ajouté le 19/02/2013
Les algues du sol en tant que participantes aux processus de formation du sol. Etude et caractérisation composition de qualité algues du sol de la division Cyanophyta. La structure et la reproduction des algues bleues. Comparaison de la composition qualitative et quantitative de Cyanophyta.